Cucurbit Genetics Cooperative Report 9:91-92 (article 28) 1986
Kwak, S.N. and J. Fujieda
University Farm, Faculty of Agriculture, Kyushu University, Kasuyamachi, Fukuoa, Japan.
We inherited a breeding program for combining the good flesh quality of C. moschata with the bush plant habit of C. pepo. Under a plastic-film house, bush plants have several advantages such as high-planting density, reduced shading effects and decreased labor. However, there is no germplasm for bush plant habit in C. moschata. Shifress (3) observed that in the F2 population of C. pepo, the segregation ratio of bush plants to vine plants was 3:1 in the early growth season, but 1:3 in the late season. He attributed this growth habit to the developmental reversal of dominance. Denna and Munger (1) observed a similar phenomenon in C. maxima, and reported that in C. pepo the bush gene is dominant to the vine gene during early growth and incompletely dominant during later growth. Zack and Loy (4) reported that the developmental reversal of dominance can be interpreted as a physiological response rather than a reversal of allelic dominance.
A bush-type C. pepo cv. Zucchini was crossed with a vine-type C. moschata cv. PM 143. PM 143 is a line selected from C. pepo x C. moschata and shows similar characteristics to C. moschata in morphology and interspecific-cross ability (2). Therefore, we temporarily classify ‘PM 143’ as C. moschata. The cross ‘Zucchini’ x ‘PM 143’ produced a few seeds with a folly developed embryo. The F1 plants showed partially self-fertility so we could raise 23 mature plants. The parents, F1 and F2 populations were grown in a plastic-film house and the internode length was measured.
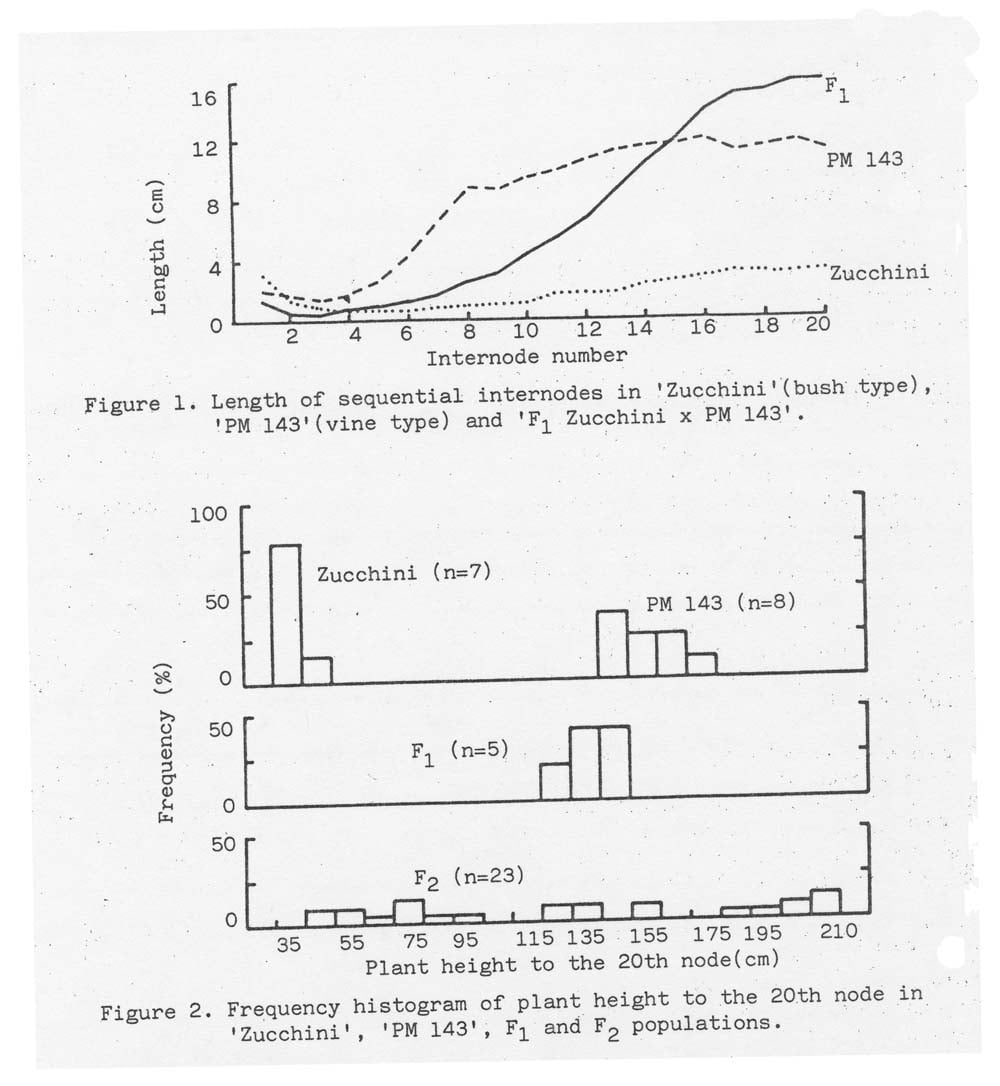
Mean length of sequential internodes of ‘Zucchini’ (bush), ‘PM 143’ (vine), and ‘F1 Zucchini x PM 143’ are presented in Figure 1. All three accessions showed a generally increased internode length although differing in length. In the F1, the first 5 internode lengths were similar to those in the bush parent ‘Zucchini’ and the internode, internode lengths of the F1 was longer than those of the vine parent ‘PM 143’. These results are similar to developmental reversal of dominance reported by other workers (1, 3).
A histogram showing the distribution of plant height to the 20th node for parental and filial accessions is presented in Figure 2. The distribution of F1 skewed toward that of the vine parent ‘PM 143’. In F2, a wide distribution with transgressive segregants (longer than 200 cm) was obtained in the plant height to the 20th node. The long internodes of F 1 plants and transgressive plants in F2 may be due to non-allelism or a major gene plus a few modifiers in C. pepo and C. moschata, or the possible influence of hybrid vigor. In the F2 population, plants displaying the bush-type habit were recovered.
These results indicate that it is possible to obtain a bush-type C moschata. Further studies are under progress for investigating detail inheritance mode of growth habit using BC, F1, F2 and F3 populations.
Literature Cited
- Denna, D.W. and H.M. Munger. 1963. Morphology of the bush and vine habits and the allelism of the bush genes in Cucurbita maxima and C. pepo squash. Proc. Amer. Soc. Hort. Sci. 82:370-377.
- Kwak, S.N. and K. Fujieda. 1985. Pollen tube growth and embryo development in interspecific crosses of Cucurbita. J. Fac. Agri., Kyushu Univ. (Japan). 30:1-8.
- Shifriss, O. 1947. Developmental reversal of dominance in Cucurbita pepo. Proc, Amer. Soc. Hort. Sci. 50:330-346.
- Zack, C.D. and J.B. Loy. 1979. The effect of light and fruit development on internode length in Cucurbita maxima squash. CGC Report 2:40-41.