Cucurbit Genetics Cooperative Report 11:60-63 (article 26) 1988
Wang Ming, Zghang Xingping and Zhang Xian
Department of Horticulture, Northwestern University, China
Na Kechi, Zhang Shuai and Zhang Juenlian
Department of Horticulture, Gansu Agricultural University, China
The development of autotriploid watermelon was a great advance in the field of watermelon breeding. However, some disadvantages still existed with this type of seedless watermelon. Partial sterility may be induced in diploid watermelon via chromosome reciprocal translocation. We used gamma-rays to irradiate the seeds of homozygous translocation strains with one translocation ring composed of 4 chromosomes (symbol (4) ). Watermelon strains were ‘Asahi Yamato’, ‘Mioyaka’, and ‘Fumin’ saent to us by H. Kihara in 1977. In order to further induce multiple reciprocal translocations for developing new few-seed/seedless watermelon strains, the seeds of the above 3 strains were sown for further selfing in 1978. The seeds of each selfed fruit were grown as a single plant line in 1979 for evaluation of their characters. In addition, some crosses between common diploid watermelon cultivars and translocations were carried out to test the seed setting rate of the heterozygous translocation strains. Some of the crosses were ‘Sugar Baby’ x ‘Asahi Yamato AT-1’ and ‘Akakotama’ x Asahi Yamato AT-2′. The plump seed setting rate of the F1 of these crosses were ca. 50%.
In general, the common cultivars of ‘Sugar Baby’, ‘Asahi Yamato’ and ‘Akakotama’ without any translocation produce ca, 500 seeds per fruit, whereas the heterozygous translocation strains contain ca. 200 seeds per fruit. Thus, they must be semi-sterile with one translocation ring at their 1st meiotic division in PMCs. It follows that the initial translocation strains were really homozygous ones with two pairs of chromosomes. In order to increase translocation chromosomes numbers/sings and decrease seed setting rate, in 1981 we used60 Co gamma rays to irradiate the seeds of the initial homozygous translocation strain “asahi Yamato.”. The does of irradiation was 50 Kr and the dose rate was 108r/min. Cytological examination indicated that some extra-ordinarily complicated aberrations of chromosome divisions occurred, such as chromosome segments, bridges, lagging chromosomes, chromosome adhesion, micronuclei, etc. However, these kinds of abnormal chromosome divisions were of no use to our breeding program, Some new useful translocations were also obtained in the following generations. In 1983, the seeds of these new strains were irradiated again by gamma-ray and induced more translocation strains with different translocation chromosomes/rings and then crossed with each other for syntheses of multiple reciprocal translocation complexes,. The translocation in heterozygous strains varied both in chromosome numbers and translocation positions. These can be described as follows: (4) + 9II (ring with 4 chromosomes + 9 bivalents), (4) + (4) + 7II, (4) + (6) + 6II, (4) + (8) + (6) + 4II, (10) + (4) + (4) + 2II, (12) + 5II and asynapsis, etc. The seed setting rate of some of the different translocation strains and their F1 hybrids are listed in Table 1.
Table 1. Seed setting rate of the parent strains and their F1 hybrids
Parent translocation strains |
Seed setting rate of parent (%) |
Cross combinations |
Seed setting F1 |
| 83-10 | 42.6 | 83-11 x 83-2 | 23.8 |
| 83-11 | 62.6 | 83-12 x 83-8 | 19.8 |
| 83-12 | 51.1 | 83-12 x 83-11 | 24.9 |
| 83-13 | ca.50 | 83-12 x 83-6 | 27.5 |
| 83-15 | 51.2 | 83-13 x 83-16 | 23.0 |
| 83-16 | ca.50 | 83-16 x 83-10 | 21.0 |
| 83-19 | 43.0 | 83-15 x 83-19 | 30.2 |
The results showed that the above translocation strains were really different ones with different translocation chromosomes. The pollen of partially sterile strains were abnormal in size, shape, and content as well. Some pollen grains wee empty and wrinkled without germinability. Some small white pollen and giant ones were also found in their anthers (Fig. 1).
Morphological effects of chromosome translocations occurred extensively, for instance, a seedling with one or three, even four cotyledons; and a plant densely covered with white hair and the leaves becoming curly resembling a virus infection. In a few plants, female flowers and fruits grew thickly and almost set at every node (Fig. 1); different kinds of rind color variations, e.g. a golden yellow color rind variation occurred in ‘Asahi Yamato’ translocation strain. In addition, the rind of some fruits became so hard hat one melon can support three persons standing on it.
Three multiple translocation strains of ‘Asahi Yamato’, Miyaka’ and ‘Fumin’ were also bred by Northwestern Agricultural University i 1984-1985. Some crosses between the above strains as well as between the ‘Fumin’ translocation strain and 16 different common watermelon cultivars (such as ‘Lovrin 532’, etc.)were conducted and 198 fruits were selected individually. The best crosses were ‘Fumin’ translocation strain crossed with ‘Charleston Gray’ and ‘Crimson Sweet’, respectively. The least number of seeds per fruit were only 25 and 52 seeds.
In brief, theoretically and practically, the more chromosome translocations involved, the higher the sterility. In this study, through self-pollination combined with segregation, selection and cross breeding, several new strains of few-seeded watermelon were developed. Their seed setting fate was 50-80% less than that of common watermelons. The extreme type contained only a few seeds, Some of the new strains are very promising for commercial production with high yield, good quality, early maturity and good storage ability.
Figure 1. Chromosomes and fruit of few-seed /seedless watermelon via chromosome reciprocal translocations induced by gamma radiation.
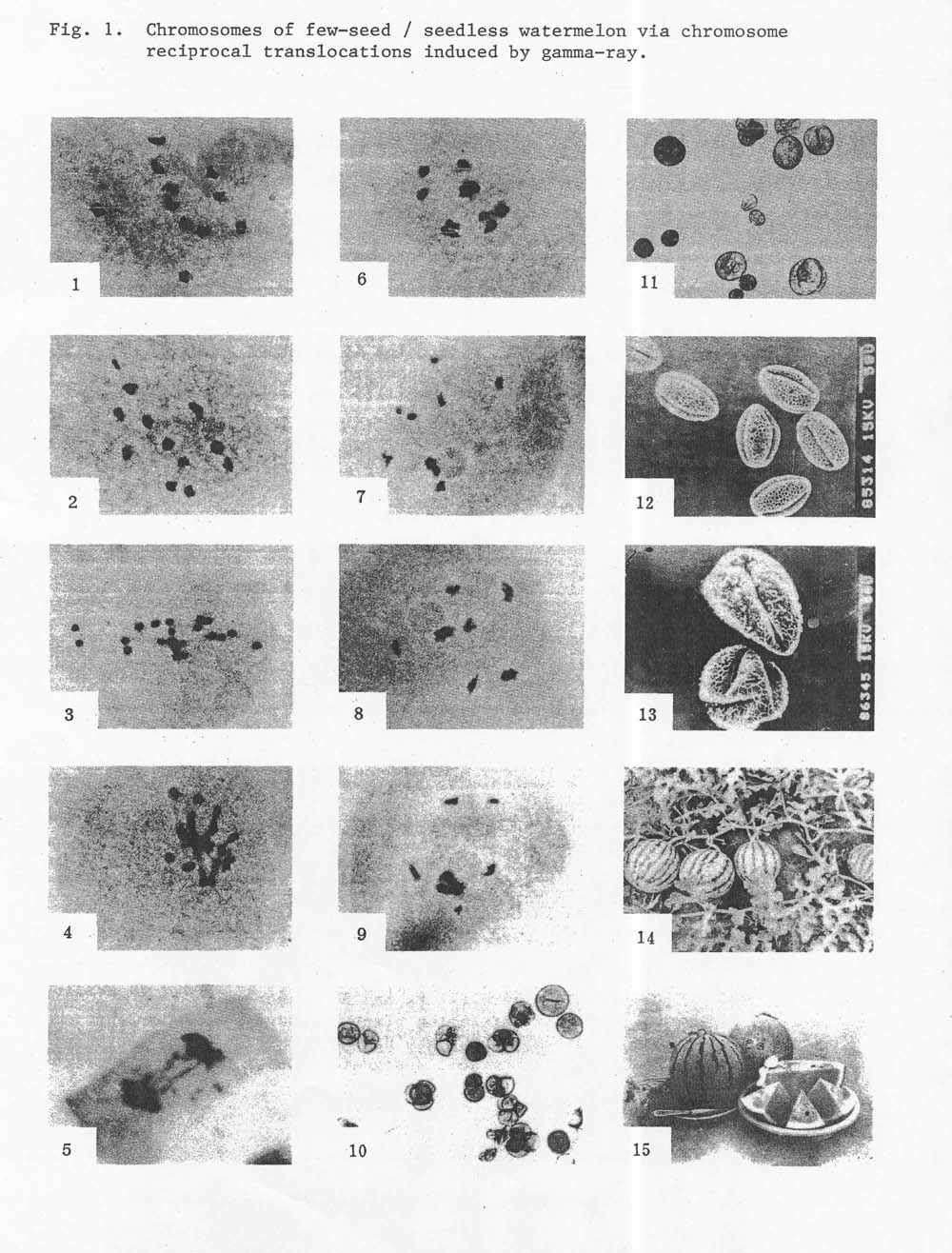
Figure 1. Chromosomes and fruit of few-seed /seedless watermelon via chromosome reciprocal translocations induced by gamma radiation. Plate 1 – normal chromosomes of watermelon: 11 bivalents (x 1000). Plate 2 – 11 bivalents + 1 segment. Plate 3 – asynapsis: 22 monovalents. Plate 4 – abnormal synapsis. Plate 5 – bridge and segments. Plate 6 – (6) + 8II (bivalents). Plate 7 – (8) + 7II. Plate 8 – (6) + (4) + (4) + 4II. Plate 9 – (12) + 5II. Plates 10 + 11 – the pollen grains of partial sterile strains showed significantly abnormal in size, shape and content., some of them adhering together, Plate 12 – the scanning electron microphoto of pollen grains: normal fertile pollen grains. Plate 13 – the scanning electron microphoto of pollen grains: sterile pollen grains, empty and wrinkled without germinability. Plate 14 – one of the very special variations: the fruits grew thickly, and all the fruits can grow to maturity for commercial use. Plate 15 – the fruits of few-seed watermelon (heterozygous translocation strains).
Literature Cited
- Hiroyuki Oka, Tadahiro Watanabe and Ichizo Nishyama. 1967. Can. J. Genet. Cytol. 9:482-489.
- Kihara, J. 1951. J. Amer. Soc. Hort. Sci. 58:217-230.
- Kihara, H., K. Salto and M. Shimotsuma. 1972. Seiken Ziho 25:63-65.
- Plant Breeding Abstracts 43(8):6384
- Katinjar, R.S. and S.K, Roy. 1980. Gene. Abs. 1981. 13(5):5062.
- Minoru Shimotsuma. 1968. Seiken Ziho 20:47-53.
- Susumu Saka, Nashimura, Honehachi. 1969. Japan. Agricultural Research Quarterly 4(3):18-21.
- Yoshinohu Egawa and Masatake Tanaka. 1984. Japan. J. Breed. 34(4):445-450.
(Chinese references omitted)