Cucurbit Genetics Cooperative Report 15:4-6 (article 2) 1992
J.E. Staub and L.D. Knerr
Vegetable Crops Research, USDA/ARS, Department of Horticulture, University of Wisconsin, Madison, WI 53706
Phylogenetic models of the genus Cucumis have been constructed based on variation in morphology and geography, cucurbitacins, flavonoids, cytological studies and chromosome banding patterns, cross-compatibility, isozymes, and nuclear DNA. The genus Cucumis has been partitioned into two subgenera (3): Cucumis (x=7) and Melo (X=12). The majority of the Cucumis species have been placed in the subgenus Melo which are indigenous mainly to Africa. Although it has been implied that the African Cucumis species arose from a common ancestor, phylogenetic relationships among three groups (annual, perennial diploid monoecious, and polyploid dioecious and monoecious) of African Cucumis species are not conclusive.
Of the four groups which comprise the subgenus Melo (3), Group 2 has recently been further subdivided into 2 subgroups using meiotic and crossability analysis, electrofusing of peroxidases, cucurbitacins, flavonoids, and geographic distribution (6). The Myriocarpus subgroup consists of C. africanus Lin. f.., C. hepadactylus Naud.(4x), C. myriocarpus Naud. ssp. leptodermis (Schweik.) Jeffrey and Halliday, and ssp. myriocarpus. The Anguria subgroup consists of C. aculeatus Cogn. (4x), C. anguria var. anguria L., C. anguria var. longipes Meeuse C. prophetarum L., and C. zeyheri Sond. (2x and 4x).
We have used isozyme analysis to describe biochemical relationships among the wild cross-compatible African diploid species of Cucumis (8). We proposed that phylogenetic affinities exist between C. anguria var. anguria, C. africanus, and C. dipsaceus, and that the relationships among C. anguria var. longipes, C. metuliferus, and C. myriocarpus are closer than had previously been proposed. In contrast, C. myriocarpus is more differentiated from C. anguria var. anguria than either C. africanus or C. dipsaceus (1, 8). Singh and Yadava (7) have argued that C. myriocarpus is genetically separated from C. dipsaceus.
Recently, we have analyzed electrophoretic variation of seven African species within Cucumis, subgenus Melo, subgroup Myriocarpus [4x C. aculeatus, C. Anguria var. anguria, C. dipsaceus, C. Ficifolius, C. Myriocarapus ssp. leptodermis Schweik., C. prophetarium, and 2x and 4x C. zeyheri Sond.], and 2x C. sativus L. of subgenus Cucumis (9). Analysis of allelic frequencies among 14 polymorphic loci indicated that C. ficifolius, C. anguria var. anguria, and 2x and 4x C. zeyheri have biochemical affinities and could be distinguished from C. aculeatus and C. dipsaceus which were similar. Within the first group, C. myriocarpus ssp. leptodermis and one C. prophetarum accession formed a well-defined group. C. savitus was dissimilar from all other species studied.
The wild African species of Cucumis are undoubtedly linked by common evolutionary episodes. Accurate phylogenetic construction requires synthesis of revel ant data and the prudent refinement of existing hypotheses. A systematic survey of all available data was conducted to develop a testable evolutionary hypothesis which might then lead to a more definitive phylogenetic construction of the wild African species (Fig. 1). From this synthesis it is hypothesized that four annual (C. africanus, C. anguria, C. dipsaceus, C. myriocarpus) and four perennial (C. aculeatus, C. ficifolius, C. figarei, C. prophetarum) species are derived from a common ancestor (Fig. 2). C. Zeyheri could be either a potential, albeit distant, progenitor species of those two groups or a derivative of a tetraploid hybrid between the two groups. Ancestral forms of 4x aculeatus or C. dipsaceus may also have provided a secondary bridge for gene transfer during the evolution of those two groups.
There are several corollaries to that hypothesis which could be proffered (Fig. , 2). For instance, since F1 hybrids between C. anguria and C. prophetarium, and between C. zeyheri and C. Prophetarum are moderately to sparingly cross-fertile (4), it seems reasonable to hypothesize that the perennial C. prophetarum may have evolved from the annual C. anguria or perennial C. zeyheri. Likewise, our results and those of Dane (2) and Nijs and Visser (4) suggest that C. ficifoloius is more reproductively isolated than C. zeyheri from all annual species except C. dipsaceus. C. ficifolius may therefore have initiated speculation earlier and/or contributed to the evolution of C. dipsaceus.
Those and other corollaries of the above working hypotheses can be tested using molecular techniques. In the study of Perl-Treves and Galun (5) the degree of homoplasy, the limited number of accessions used, and the number of “less characterized” and back or parallel “point mutations” did not allow them to clearly establish relationships among African species which formed a compact group. A larger number of accessions and endonucleases could be utilized to construct a rooted parsimonious tree. The working hypotheses presented above could then be tested using specific outgroups to make synaptomorphic comparisons.
Figure 1. A composite diagram of cytogenetic, biochemical and cross-compatibility relationships among eight African Cucumis species. Weak affinities (a synthesis of all criteria) of C. zeyheri 4x with other species is represented by a rectangle.
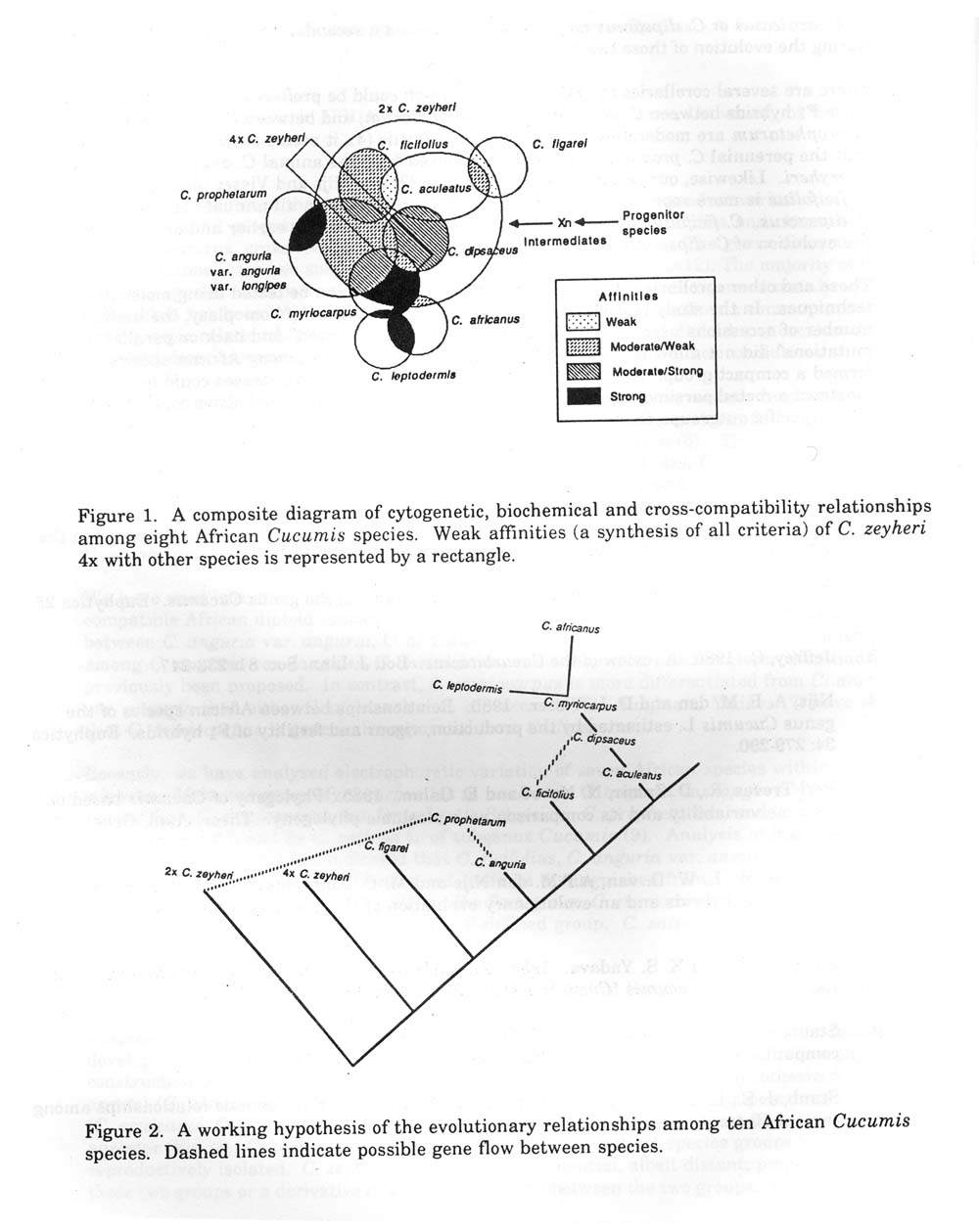
Figure 2. A working hypothesis of the evoluationary relationships among ten African Cucumis species. Dashed lines indicate possible gene flow between species.
Literature Cited
- Dane, F., D.W. Denna and T. Tsuchiua. 1980. Evolutionary studies of wild species in the genus Cucumis. Z. Pflanzenzuchtg. 85: 89-109.
- Dane, F. and Tsuchiya, T. 1976. Chromosome studies int he genus Cucumis. Euphytica 25: 367-374.
- Jeffrey, C. 1980. A review of the Cucurbitaceae. Bot. J. Linn. Soc. 81: 233-247.
- Nijs, A.P.M. den and D.L. Visser. 1985. Relationships between African species of the genus Cucumis L. estimated by the production, vigour and fertility of F1 hybrids. Euphytica 34: 279-290.
- Perl-Treves, R., D. Zamir, N. Navot and E. Galun. 1985. Phylogeny of Cucumis based on isozyme variability and its comparison with plastome phylogeny. Theor. Appl. Genet. 71: 430-436.
- Raamsdonk, L.W D van, A.P.M. den Nikjs and M.C. Jongerius. 1989. Meiotic analyses of Cucumis hybrids and an evolutionary evaluation of the genus Cucumis. Plt. Sys. Evol. 163: 133-146.
- Singh, A.K. and K.S. Yadava. 1984. An analysis of interspecific hybrids and phylogenetic implications in Cucumis (Cucurbitaceae). Plant Syst, Evol. 147: 237-252.
- Staub, J.E., L. Frederick and T.L. Marty. 1987. Electrophoretic variation in cross-compatible wild diploid species of Cucumis. Can. J. Bot. 65: 792-798.
- Staub, J.E., L.D. Knerr, D.J. Holder and B. May. 1992. Phylogenetic relationships among several Cucumis species. Can. J. Bot. (in press).