Cucurbit Genetics Cooperative Report 16:84-86 (article 32) 1993
M. Abdul Vahab and K.V. Peter
College of Horticulture, Vellanikkara, India – 680 654
Bittergourd (Momordica charantia) is an important fruit vegetable in the warm humid tropics, grown throughout the year. Preference for this vegetable varies highly with the extent of bitterness. In the present study an attempt was made to cross M. charantia with M diooica, the bitterless, small fruited, tuberous perennial to seekpossibilities of transferring desirable attributes of the latter to the former.
Both direct and reciprocal crosses were made between M. charantia and M. dioica. Anthesis in M. charantia is between 4:00 and 10:00 a.m. while that in M. dioica is between 5:30 a.m. and 12:00 p.m. Therefore, pollen from both species were collected at anthesis time, stored at 10 ˚ C and hand pollinated at the corresponding time of anthesis in both species.
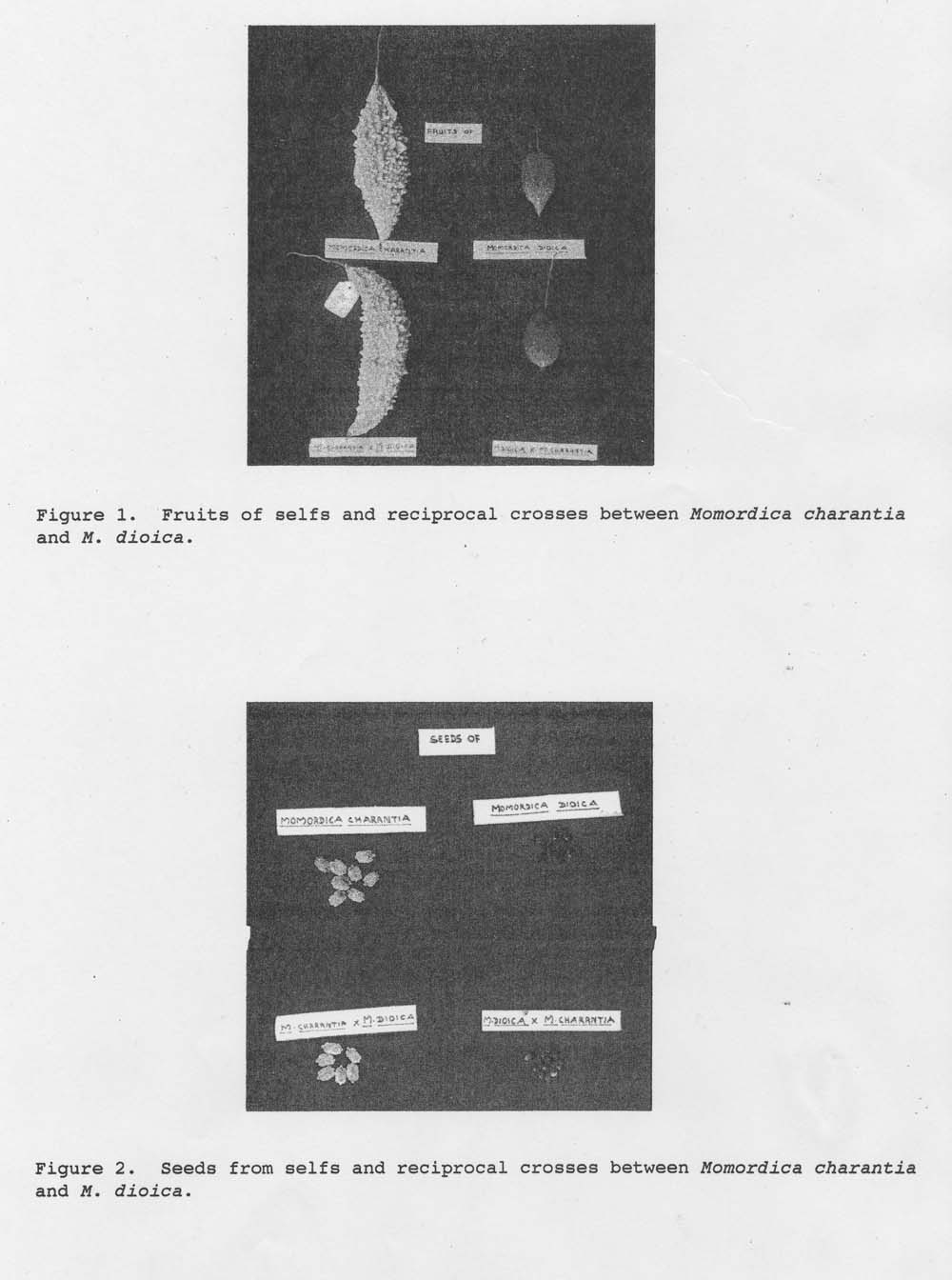
The results (Table 1) indicated that the cross M. charantia x M. dioica or its reciprocal failed to set fruit when normal pollens were used as reporte earlier (Trivedi and Roy, 1972; Dutt and Pandey, 1982). The ovary did not grow further; it shriveled and dried in three days. However, when crosses were made using pollens collected and stored at 10 ˚ C, the percentage of success was above 90. The fruits and F1 seeds resembled their respective female parents in both direct and reciprocal crosses (Fig. 1 and 2). The number of chaffy seeds was more in crossed fruits than in selfed fruits. Further studies are in progress.
This is the first report of success of crossing between M. charantia and M. dioica. The study indicated the possibility of utilizing the bitterless, perennial tuberous M. dioica in transferring the desirable attributes to the commercially cultivated large fruited bittergourd. The study also shows the possibility of identifying the progenitor of bittergourd through genome analysis. the failure in earlier studies could be attributed to nonsynchronization of anthesis in two species, which is solved through storing pollens at low temperature and pollinating at anthesis in both species.
Table 1. Crossability between M. charantia and M. dioica.
Number of |
|||||
Crosses/selfs |
|||||
Crosses or self |
Fruit set |
Seeds/fruit |
Bold seeds |
Chaffy seeds |
|
| M. charantia x M. dioica | 50 | 46 | 23 | 14 | 9 |
| (using pollens stored at 10 ˚ C) | |||||
| M. dioica x M. charantia | 50 | 47 | 21 | 11 | 10 |
| (using pollens stored at 10 ˚ C) | |||||
| M. charantia | 25 | 24 | 22 | 19 | 3 |
| (selfed) | |||||
| M. dioica | 25 | 23 | 20 | 18 | 2 |
| (selfed) | |||||
| M. charantia x M. dioica | 50 | 0 | — | — | — |
| (using normally collected pollens) | |||||
| M. dioica x M. charantia | 50 | 0 | — | — | — |
| (using normally collected pollens) | |||||
Figure 1. Fruits of selfs and reciprocal crosses between Momordica charantia and M. dioica.
Figure 2. Seeds from selfs and reciprocal crosses between Momordica charantia nd M. dioica.
Literature Cited
- Dutt, B. and C. Pandey. 1982. Callose deposition in relation to incompatibility in Momordica In: Roy, R,P. and U. Sinha (Ed.) Cytogenetics in India.
- Trivedi, R.N. and P.P. Roy. 1972. Cytological studies insome species of Momordica. Genetica 43:282-291.