Cucurbit Genetics Cooperative Report 20:11-12 (article 6) 1997
S. Serce and J.E. Staub
Vegetable Crops Research Unit, USDA/ARS, Departments of Horticulture, University of Wisconsin-Madison, WI. 53706 U.S.A.
Introduction. Cucumber vine growth, lateral branching, leaf area, fruit set, and growth rate are adversely affected when plants are subjected to soil moisture deficits (Cummins and Kretchman, 1975). Fruit yield and quality are also affect by plant water stress (Navazio and Staub, 1994; Staub et al., 1988; Thomas and Staub, 1992).
Cultivar differences in fruit quality have been observed in responses to water stress (Navaho and Stub, 1994; Stub et al., 1988). The plant response of different genotypes depends on the stress environment (Navaho and Stub, 1994). Moreover, although a plant may not exhibit symptoms of stress during periods of soil moisture deficits, it is possible that physiological responses may occur during incipient stress and cause poor fruit quality. The hypotheses as to cause and effect, and genetic background could be tested using nearly-isogenic lines differing in leaf size and plant architecture. Here we report the effects of water stress on seed size and fruit cavity size in nearly-isogenic lines differing plant habits to assess the role of genetic background during water stress.
Material and Methods. Three nearly-isogenic lines (LLDeDe = normal leaf, indeterminate; Ildede = little leaf, determinate; llDeDe = little leaf, indeterminate) were compared for response to soil moisture deficits in the summer of 1996. Nearly-isogenic lines (BC6S3) received either adequate water during fruit enlargement (well-watered; control) or were subjected to low soil moisture tensions (water-withheld; stress).
The experiment was constructed at the University of Wisconsin Research Station, Hancock, Wis, on a plainfield loamy sand, typic updisamment; mixed mesic. Plants were arranged in adjacent, randomly-assigned treatment plots (control and water withheld) 4.0 m in apart. Each plot was divided into four 6.0 m rows, with end (1 m) and side (8 m) plot borders. Cucumbers was planted on 1.5 row centers and spaced approximately 10 cm apart in the row (67,000 plants/ha). Each treatment row was divided into 1 m segments and genotypes was randomized in each of five replications. Standard fertilization, spraying, and hand cultivation were practiced in all experiments.
After harvest (when 5% of the fruit were 5 cm in diameter) and postharvest storage, fruits were fresh-pack processed. Processed cucumbers were held for 12 to 14 weeks at 8 C prior to evaluation.
All fruits were halved in longitudinal section. Seed size and fruit cavity (endocarp) size of samplers were evaluated by eight people. Theses fruit characters were rated using 1 to 4 scale (1 = small seed and cavity size; 4 = large seed and cavity size). ANOVA and mean separation of evaluation seed development, and cavity size were performed separately using a split-split-split-plot analysis. Mean separations for all factors in all ANOVAs were performed with Fisher’s protected LSD at the 5% level (Steele and Torrie, 1980).
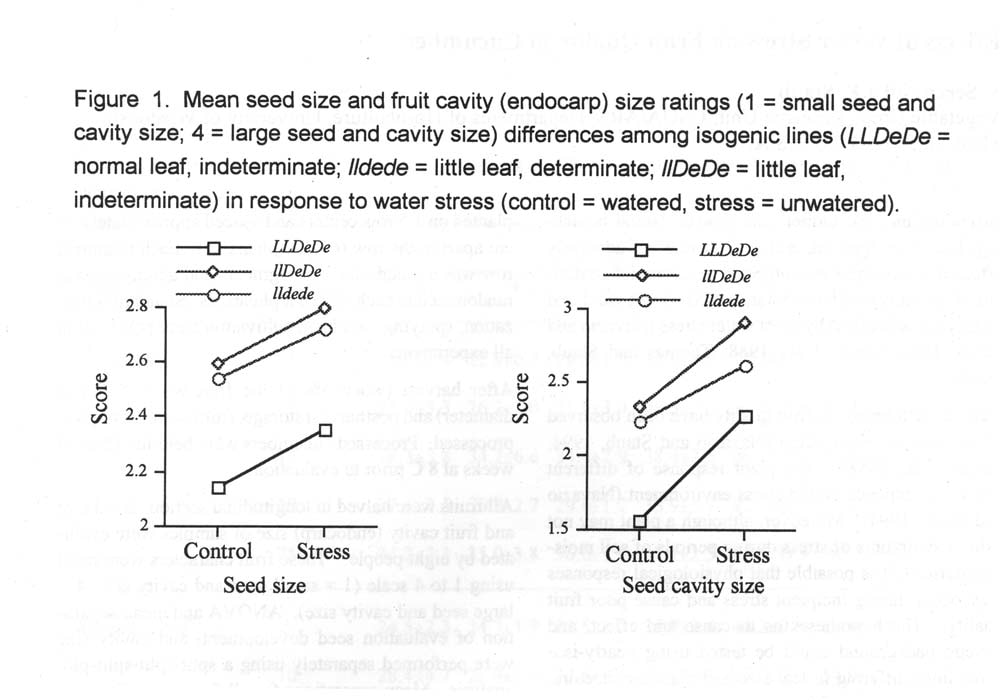
Results and Discussion. Differences were observed among nearly-isogenic lines for seed size and seed cavity size (Figure 1). Although plant habit (de vs. De) had no affect on the fruit parameters evaluated, leaf size (ll vs. LL) did. The normal leaf line had significantly smaller seeds and cavity size when compare to its little leaf counter part either in determinate or indeterminate backgrounds. This suggests that seed in little leaf lines were maturing more rapidly than those in the normal leaf line. This accelerated maturation in fruit of little leaf lines was also evidenced by the presence of larger seed cavities in these fruit. Both of these observations are indicative of a response to water stress imposed by a soil moisture deficit. This response in little leaf germplasm is manifested by more rapid senescence (i.e., fruit maturation) when compared to well-watered controls and normal leaf germplasm.
Navazio and Staub (1994) observed that although normal leaf cultivars wilted when subjected to short periods of soil moisture deficits, little leaf lines did not. This result was confirmed in our study (data not presented). Thus, our results suggest that enlarging fruit of little leaf genotypes can be subjected to stress, even though the plant is not showing wilt symptoms.
Figure 1. Mean seed size and fruit cavity (endocarp) size rating (1 = small seed and cavity size; 4 = large seed and cavity size) differences among isogenic lines (LLDeDe = normal leaf, indeterminate; lldede = little leaf, determinate; llDeDe = little leaf, indeterminate) in response to water stress (control = watered, stress = unwatered).
Literature Cited
- Cummins, T.C. and D.W. Kretchman. 1975. Relation of internal water to growth and development of the picking cucumber. HortScience 10:319. (Abstr.)
- Navazio, J.P. and J.E. Staub. 1994. Effects of soil moisture, cultivar, and postharvest handling on pillowy fruit disorder in cucumber. J. Amer. Soc. Hort. Sci. 119:1234-1242.
- Staub, J.E., P. Rousos, and B.E. Struckmeyer. 1988. Anatomical characterization and possible role of calcium in “pillowy,” a fruit disorder in processing cucumber. J. Amer. Soc. Hort. Sci. 113:905-909.
- Steel, R. and J.H. Torrie. 1980. Principles and procedures of statistics. 2nd ed. McGraw-Hill, New York.
- Thomas, R.S. and J.E. Staub. 1992. Water stress and storage environment affect pillowy fruit disorder in cucumber. J. Amer. Soc. Hort. Sci. 117:394-399.