Cucurbit Genetics Cooperative Report 23:117-119 (article 39) 2000
Tamas Lelley and Silvia Hengmuller
Department of Biotechnology in Plant Production, Institute of Agrobiotecnology, A-3430 Tuln, Austria, Konrad Lorenz Str. 20
Abstract: For the first time in Austria, the year 1997 brought a severe infection of oil pumpkin by the Zucchini Yellow Mosaic Virus (ZYMV) causing a 50% loss in yield. an initiative to introduce resistance into Austrian germplasm by breeding found immediate acceptance and financial support by the Federal Ministry of Agriculture and Forestry and by the primarily affected states Styria, Burgenland and Lower-Austria.
Since, Hitherto, no resistance or tolerance genes were found in the species cucurbita pepo , genes of tolerance originally derived from Cucurbita moschata are being introduced in Austrian germplasm via crosses with Zucchini varieties possessing such tolerance. Sincere tolerance in this material appears to be a recessive trait, to aid selection attempts are being made to find a molecular marker linked to the tolerance gene, using bulked segregant analysis (BSA). F2 derived F3 plants were tested with the virus isolated in austria to determine the genetic constitution of the F2 plants for creating the DNA bulks parallel to the BC2F1 plants were produced. If BSA will be successful in the winter 99/00, plants having the tolerance gene in heterozygous condition inBC2F1 plants can be selected for selfing or further backcrossing.
Keywords: ZYMV, Zucchini Yellow Mosaik Virus, molecular marker, RAPD, BSA, Bulked Segregant Analysis
Introduction: Zucchini yellow mosaic virus (ZYMV) is one of the most destructive pathogens infecting cucurbits including oilseed pumpkin. The virus was first described in Italy 1973 (1,2); later several ep9idemics were reported (3,4). In C. pepo or its close wild relatives no resistance to this virus has been found so far (5); however, resistance is available in some accessions of C. moschata from Nigeria and Portugal. It was described as a single incompletely dominant gene, Zym (6,7). The only wild species up to now identified as carrying resistance to ZYMV is C. ecuadorensis. This resistance is also conferred by a single, incompletely dominant gene, Zym (8). Direct transfer via interspecific crosses can only be expected to be successful in C. maxima (9).
Traditionally, a specific “hull-less” or “naked-seeded” pumpkin cultivar C. pepo var. styriaca has been grown in Austria for seed-oil production for well over 100years.In 1997, for the first time, a severe ZYMV epidemic destroyed about 50% of the pumpkin harvest. This happened at a time when increasing consumer awareness for taste, nutritive value and potential health effects of pumpkin seed oil caused an expansion of the growing area. An initiative to introduce resistance into austrian germplasm by breeding found immediate acceptance and financial support by the Federal Ministry of Agriculture and Forestry and by the most highly affected states Styria,Burgenland, and Lower-Austria.
This presentation describes the progress of this work, starting from its inception in the year 1998.
The breeding strategy: An extensive search for sources of resistance led to the identification of several zucchini varieties in the U.S.A. which possess a gene for tolerance against ZYMV. This gene was first reported in a genotype from Nigeria (“Nigerian Local”) and, based on crossing experiments with other C. moschata cultivars, has been described as being partially dominant as a result of having some modifier genes for a single resistance gene (6). Transferring this gene to C. pepo summer squashes improved their tolerance to ZYMV, however, not to the same extent as in C. moschata (6). Nevertheless, a limited number of zucchini cultivars possessing this gene have been released by a few American seed companies.
Seeds of the varieties Jaguar, Tigress, and Puma were obtained from the Harris Moran Seed Company and of the varieties Dividend and Revenue from Novartis U.S.A. They were described as highly tolerant against isolates of ZYMV from America and China (10). All these genotypes were tested, together with Austrian breeding lines, against the virus isolated in Austria. This experiment has proven the good tolerance of the zucchini varieties from America when compared to the high susceptibility of the Austrian material. An artificial infection of the two cotyledons and the first leaf with a high virus titer of the inoculum, while more or less killing the Austrian genotypes (strong mottling, deep foliar serration of young leaves, stunting or death), caused slight leaf symptoms, no stunting and good fruit formation, although protrusions could be observed on many of the fruits. Crosses of these varieties with Austrian germplasm were successful. The F1 was again tested with the Austrian isolate of the virus. All F1 combinations were partly selfed, partly back-crossed with the respective Austrian parents.
Later F2 progenies of two combinations were selected and tested with the virus. One has shown a 37:11 susceptibility to tolerant segregation, the other combination segregated 41:7. Testing 10 to 15 F3 progenies of 48 and 47 selfed F2 plants of the two selected crosses identified highly susceptible and tolerant F2 progenies. The fact that in the tolerant progenies a few highly susceptible plants regularly occurred, suggests that either tolerance is determined not only by one single gene, or that genetic background plays a role in the expression of the character.
DNA of tolerant plants of each of 12 different tolerant F2 progenies and from susceptible single plants of each of 12 different highly susceptible F2 progenies were collected. DNA will be pooled separately for a bulked segregant analysis.
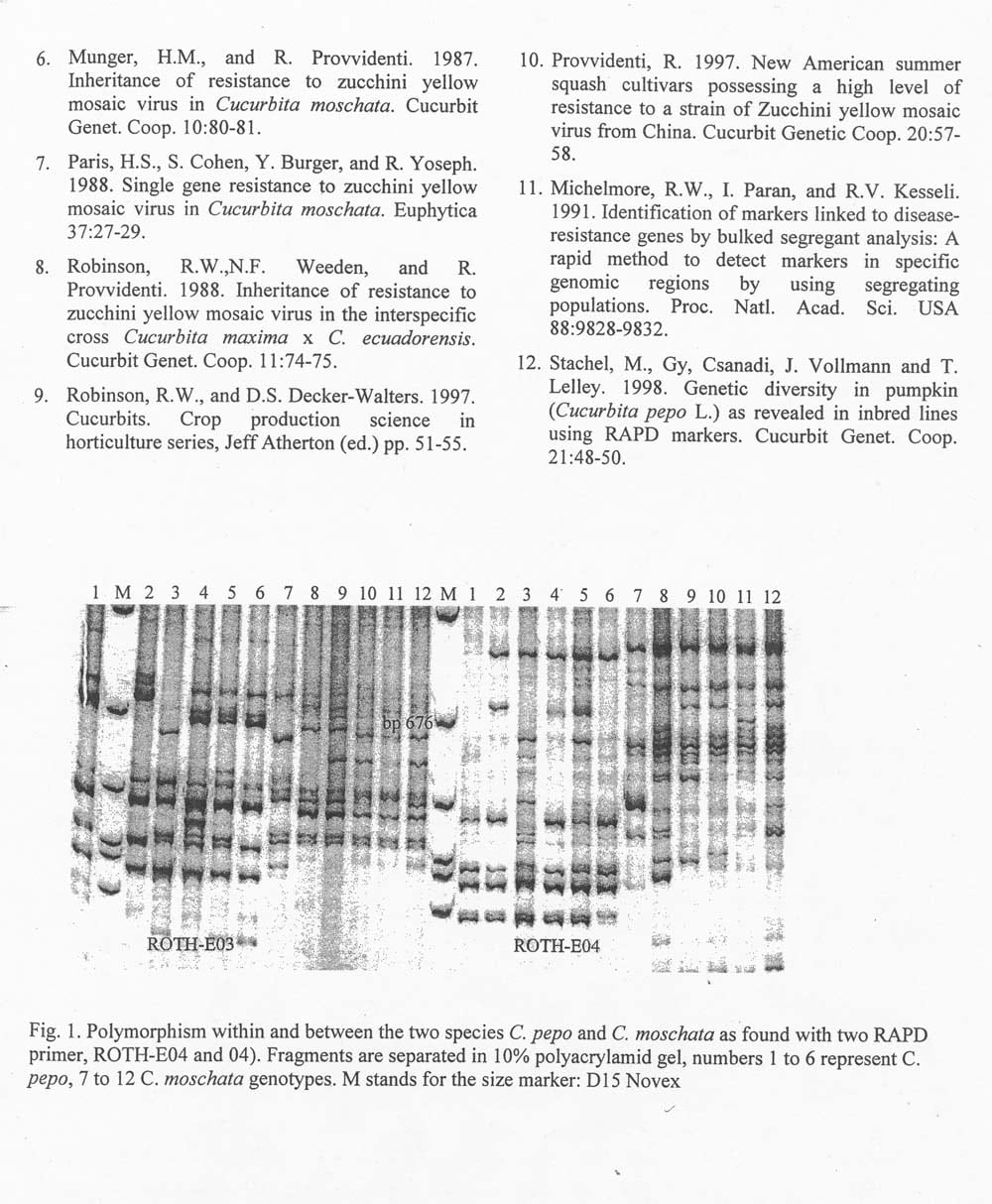
The DNA approach: Bulked segregant analysis (11)m i.e. testing two DNA-pools with isogenic background differing only in one phenotypic character for polymorphism, is a very efficient method for finding a molecular market closely linked to a gene determining this character. Prerequisite is clear distinction of genotype classes, susceptible versus tolerant, and sufficient polymorphism between donor (C. moschata) and receptor (C. pepo) at the DNA level. The method is especially useful for dominant type markers i.e. RAPD or AFLP. Analyzing genetic relatedness of different pumpkin inbred lines revealed a high level of polymorphism within C. pepo using RAPD marker (12). Even more polymorphism can be expected between the two species (Fig. 1). Therefore, it can be expected that DNA polymorphism between these two species, close enoughto the gene for tolerance, will be found when a sufficient number of markers has been tested. At present, F2 derived F3 populations have already been tested for tolerance or high susceptibility and were identified. DNA from these plants has been isolated and isogenic pools established. Testing of markers for polymorphism on these two pools is in progress.
Acknowledgement. All virus testing is being carried out in the Federal Office and Research Center for Agriculture under the supervision of DIU M. Riedle-Bauer, who also performs the grading of infection. In this connection special thanks are due to Ms. Betty Suarez for her outstanding technical assistance. This project is financially supported by the Federal Ministry of Agriculture and Forestry, by the States Styria, Burgenland and Lower-Austria and by the Breeding Company Gleisdorfer Saatzucht.
Literature Cited
- Lecoq, H., M. Pitra, and M. Clement. 1981. Identification et caracterisation d’un potyvirus provoquant la maladie due rabougrissement jaune du melon. Agronomie 1:827-834.
- Lisa, C., G. Boccardo, D’Agostino, G. Dellavalle, andM. D’Aquino. 1981. Characterization of a potyvirus that causes zucchini yellow mosaic. Phytopathology 71:668-672.
- Provvidenti, R. H.M. Munger, and A.O. Paulus. 1984. Epidemics of zucchini yellow mosaic virus and other cucurbit viruses in Egypt in the spring of 1983. Cucurbit Genetic Coop. 7:78-79.
- Provvidenti, R. 1986. Occurrence of Zucchini yellow mosaic virus in the United States in 1985. Cucurbit Genetic Coop. 9-96.
- Provvidenti, R. 1993. Resistance to viral diseases of cucurbits. In: Resistance to viral diseases of vegetables: genetics and Breeding. Molly M. Kyle (ed). Timber Press, Portland Oregon. 8-43.
- Munger, H.M., and R. Provvidenti. 1987. Inheritance of resistance to zucchini yellow mosaic virus in Cucurbita moschata. Cucurbit Genet. Coop. 10:80-81.
- Paris, H.S., S. Cohen, Y. Burger, and R. Yoseph. 1988. Single gene resistance to zucchini yellow mosaic virus in Cucurbita moschata. Euphytica 37:27-29.
- Robinson, R.W., N.F. Weeden, and R. Provvidenti. 1988. Inheritance of resistance to zucchini yellow mosaic virus in the interspecific cross Cucurbita maxima x C. ecuadorensis. Cucurbit Genet. Coop. 11:74-75.
- Robinson, R.W., and D.S. Decker-Walters. 1997. Cucurbita. Crop production science in horticulture series, Jeff Atherton (ed.) pp. 51-55.
- Provvidenti, R. 1997. New American summer squash cultivars possessing a high level of resistance to a strain of Zucchini yellow mosaic virus from China. Cucurbit Genetic Coop. 20:57-58.
- Michelmore, R.W., I. Paran, and R.V. Kesseli. 1991. Identification of markers linked to disease-resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations. Proc. Natl. Acad. Sci. USA 88:9828-9832.
- Stachel, M., Gy, Csanadi, J. Vollman and T. Lelley. 1998. Genetic diversity in pumpkin (Cucurbita pepo L.) as revealed in inbred lines using RAPD markers. Cucurbit Genet. Coop. 21:48-50.