Cucurbit Genetics Cooperative Report 23:41-45 (article 12) 2000
J.G. de A. Assis, A.L.P.C. de Oliveria
Instituto de Biologia, Universidade Federal da Bahia, Salvador, Bahia, Brazil, 40.170-110
A.R. Lima, I.C. Crepaldi, J.R.F. de Santana
Departmento de Biologia, Universidade Estadual de Feira de Santana, Feira de Santana, Bahia, Brazil
Introduction. Life cycles and the reproductive biology of plants are fundamental in determining cultivation and/or breeding strategies. Cucurbit species have an annual and a perennial cycle, and may have intraspecific variation in the phenological cycle. Thus, the determination of the phenological cycle may lead to the determination of genotypes which produce earlier fruits.
Various factors related to cucurbit reproductive biology have been investigated: proportion of male and female flowers, anthesis time, flowering time and duration, time of flower opening, and stigma receptivity and flower bud development (4). Studies on sex expression in some cucurbit species have shown that there are diverse phenotypes (monoecious, gynoecious, gynomonoecious and androecious). The most common type is monoecious, with the sexes in separate flowers (5).
Morphological characterization enables cultivars to be differentiated, as was shown by Wendel and Weeden (11) and Brandao et al., (1) who identified three Cucurbita pepo L. cultivars and five cucurbita maxima (Duschesne) cultivars, respectively, using seedling traits. Thirty-nine Citrullus lanatus accesses from Northeast Brazil were characterized and the genetic variability quantified using a descriptor list (7). Cucumis melo fruits are extremely polymorphic for shape and color (globe shaped, oval, elongated oblong, pubescent or glabrous, pale yellow, canary yellow or green in color).
‘Melao coalhada’ (Fig. 1) and ‘Maxixe italiano’ (Fig. 2) are commercialized on a small scale for consumption in juices and salads, respectively. Because of their potential expanded cultivation and breeding work, this study was carried out to obtain data on some of their phenological, plant, morphological and genetic (cytogenic and isoenzymatic) aspects.
Material and Methods.The plants described in this study were collected in the semi-arid area of the state of Bahia, in Baixa Grande county, where they are locally known as ‘Melao coalhada’ and ‘Maxixe italiano’. The experiment was conducted in the field at Universidade Estadual de Feira de Santant (UEFS). Twenty seed per variety were sown at 2 seeds per hole, 1.5 m plant spacing, and 2.0 m row spacing in December, 1997.
The germination period was observed in days, along with the germination percentage and cotyledon and true leaf colors. The plant development was assessed by scoring the percentage of established plants, cotyledon and first true leaf color, length of the main branch, number of side branches, flower traits and flowering time, days to harvest of the first fruit, and the harvest peak. The presence of visiting insects was recorded. The fruit were characterized for size, shape, flesh color and mean number of seeds produced. They were later stored at room temperature and at 5˚C, and the postharvest shelf-life was assessed after 15 days.
Cytogenetic analyses were carried out using the conventions Feulgen method (8) and electrophoresis in starch gels (12%) for the following enzymatic systems: esterase (EST), phosphoglucoisomerase (PGI), shikimate dehydrogenase (SKDH), malic enzyme (ME), peroxidase (PER), catalase (CAT), glutamate dehydrogenase (GDH), and glutamate oxaloacetate trasaminase (GOT). Cucumis anguriaand C. sativus varieties were used to compare.
Isoenzymatic patterns from ‘Melao coalhada’ and “Maxixe italiano’ were compared with one cantaloupe type. Cotyledon leaf tissue was used in the analyses and the migration methods were lithium-borate pH 8.3 for PGI, PER, GOT, SKDH (10), and tris-citrate pH.7.5 for CAT, EM, EST, and GDH (9).
Results and Discussion. Both the varieties germinated completely in 8 to 13 days and had a high percentage of plant establishment (Table 1).
‘Melao coalhada’ was approximately 130 cm long from the main branch while ‘Maxixe italiano’ was only 88 cm long (Table 2). Similarly, ‘Maxixe italiano’ flowers are smaller in size (Figure 3) than the commercial melon varieties. flowering occurred between 40 and 75 days after planting and the flowers of both varieties are single sex. The only visiting insects observed were Apis melifera, and the flowering periods coincided allowing cross fertilization and exchange of genotypes favorable to the crop.
‘melao coalhada’ was the earliest variety, with ripe fruit at 68 days after planting, and it had a lower number of seeds compared to ‘Maxixe italiano’ (Table 3). Further study should be done at different planting times to get a clearer picture of these traits.
‘Maxixe italiano’ has potential for large scale cultivation in the Brazilian northeast because of its organoleptic characteristics, which are similar to the cucumber (Cucumis sativus), its adaptation to semi-arid climatic conditions, and its small plant size, which makes it suitable for cultivation in smaller areas than those required by other cucurbits. Also, it maintains its fruit characteristics during a storage period (15 days) at 5˚C.
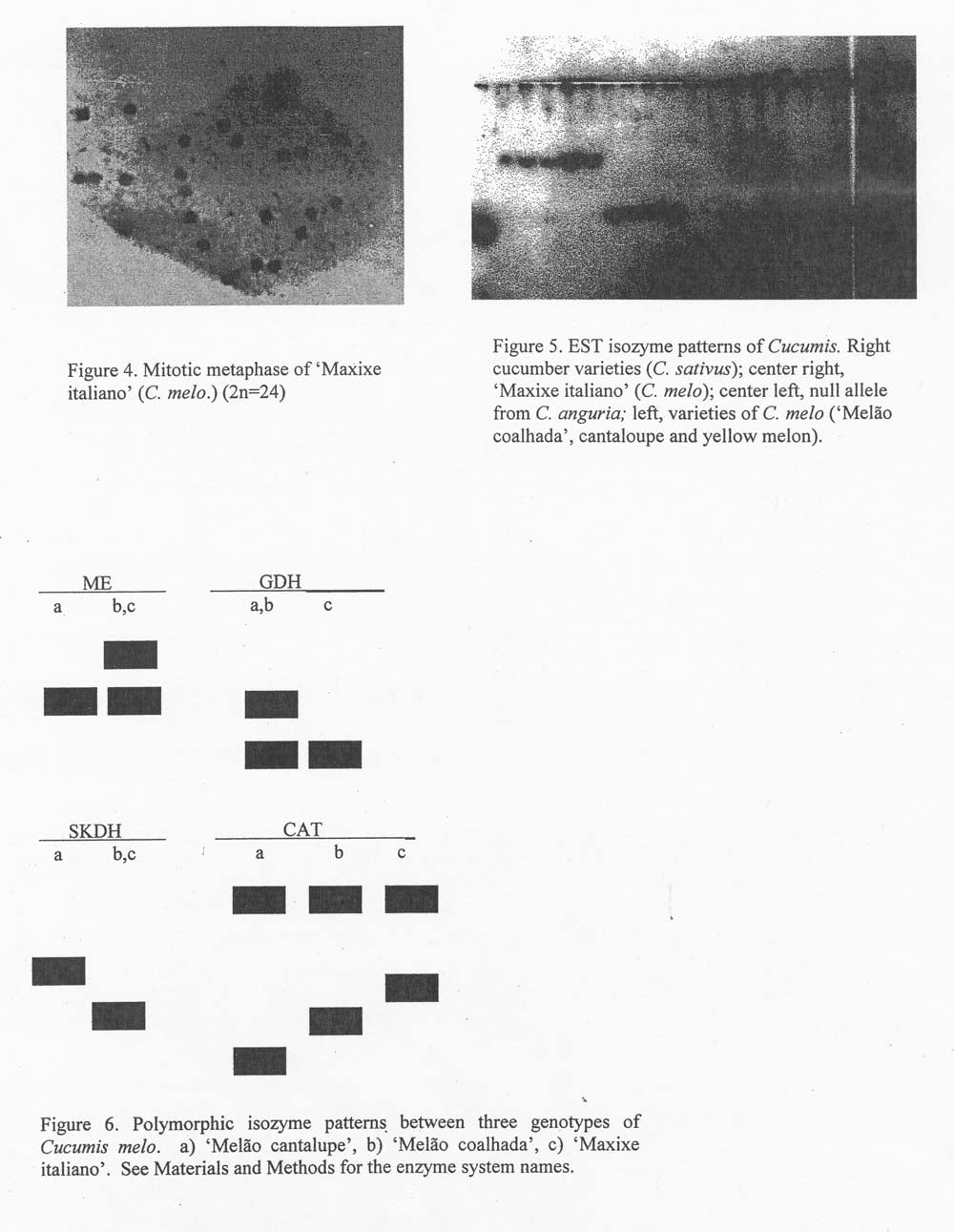
The chromosome analyses revealed a diploid number 2n = 24 for both varieties (Figure 4), as observed for the other varieties of C. melo(2, 6). Cucumis has two basic numbers: species originating in India such as the cucumber are 2n = 14, whereas species such as C. melo andC. agrarian in Africa are 2n = 24. These were introduced to Brazil by the Negro slaves, and are today widely distributed throughout the Brazilian Northeast, where they are found with great morphological diversity which is reflected in innumerable varieties.
No differences were observed at the genomic group level in any enzymatic system x = 12 (C. melo andC. anguria) and x = 7 (C. sativus). The electrophoretic patterns were monomorphic for PER, EST, GOT and PGI systems. In other systems, differences were observed between Cucumis melo, Cucumis anguria and Cucumis sativus, showing characteristic patterns for each species (Figure 5). GDH, SKDH and ME had a polymorphic locus within C. melo. CAT showed two loci, with Cat-2 being polymorphic and having three different alleles, each one observed in a different genotype (Figure 6).
Table 1. Assessment of germination and seedling stages of two local varieties of Cucumis melo.
| Variety | Germination (days) | Germination (%) | Cotyledon color | First true leave color |
| Melao coalhada | 10 | 90 | green | green |
| Maxixe italiano | 13 | 100 | green-yellow | green |
Table 2. Establishment, growth, and reproductive cycle evaluation of two local varieties of Cucumis melo
| Variety | Established plants (%) | Main branch length (cm) | Side branch No. | Days to first flower | Days to harvest of the first fruit | Days to harvest peak |
| Melao coalhada | 90 | 129.11 | 5.7 | 41 | 68 | 89 |
| Maxixe italiano | 70 | 88.28 | 4.4 | 41 | 75 | 99 |
Table 3. Fruit characteristics of two local varieties of Cucumis melo
| Variety | Fruits per plant | Fruit length (cm) | Fruit diameter (cm) | Fruit shape | Fruit weight (g) | Flesh color | Seed number |
| Melao coalhada | 1.89 | 24.11 | 22.90 | oblong | 411.61 | pale yellow | 547 |
| Maxixe italiano | 2.14 | 9.38 | 17.66 | globular | 126.60 | white | 689 |

Literature Cited
- Brandao, J.C., V.M. de M. Andrae, and M.E. S.P. Dematte. 1981. Caracterizacao de Cucurbitaceae em fase inicial de desenvolvimento 2. Comparacao de plantulas de diferentes cultivares de Cucurbita maximaDuchenese. Cientifica 9(1): 113-119.
- Dane, F. and T. Tsuchya,. 1976. Chromosome studies in the genus Cucumis. Euphytica 25:367-374.
- Esquinas – Alcazar, J.T., and P.J. Gulick. Genetic Resources of Cucurbitaceae. Rome, OBPGR, 1983. 101p. (IBPGR – 82184).
- Katrodia, J.S., P. Nath, and O.P. Dutta. 1974. Studies on floral biology in parents, F1 . F2 and backcross generations in Citrullus lanatus Thunb Mausf. Indian Journal of Horticulture 31(1):56-65.
- Lopes, J.F. 1982. Melhoramento genetico (chuchu melanicia, melao e pepino). Inf. Agropec. 8(85):61-64.
- Ramachandran, C., V.S. Seshardri, and R.A. Pai. 1985. Cytogenetical studies on dessert and non-dessert forms of muskmelon (cucumis melo L.). Cytologia 50:631-641.
- Romao, R.L. 1995. Dinamica evolutiva e variabilidade de populacoes de melancia Citrulus lanatus(Thunb.) Matsum. & Nakai em tres regioes do Nordeste brasileiro. Piracicaba. Dissertacao (Mestrado) 75p.
- Sharma, A.K, and A. Sharma. 1989. Advances in cell and chromosome research. Oxford and IBH Pub. Co., New Delhi.
- Soltis, D.E., C.H. Haufler and D.C. Darrow. 1983. Starch gel electrophoresis of ferns: a compilation of grinding buffers, gel and electrode buffers, and staining schedules. American Fern Journal 73:9-27.
- Wendel, J.P. and N.F. Weeden. 1990. Visualization and interpretation of plant isozymes. In: Soltis, D.E. and P.S. Soltis (eds.), Isozymes in plant biology. London: Chapmam and Hall p. 5-45.
- Zaccaro, R.PO., M.E.S.P. Dematte, and V.M. de M. Andrade. 1981. Caracterizaco de Cucurbitaceae em fase inicial de desenvolvimento. 3. Comparacao de plantulas de diferentes cultivares de Cucurbit pepo L. (aboboreira). Cientifica 9(2):281-287.