Cucurbit Genetics Cooperative Report 23:73-77 (article 25) 2000
Miroslav Baranek, Gergtud Stift, Johann Vollmann and Tamas Lelley
Department of Plant Biotechnology, Institute of Agrobiotechnology Tulln, A-3400 Tulln, Austria: [email protected]
Introduction. Genes not available in a species can often be obtained through interspecific hybridization. A good example is resistance against Zucchini Yellow Mosaic Virus (ZYMV), which is not found in C. pepo (6), but is available in C. moschata (3, 4, 2). Crossing the two species is difficult, though not impossible. Hybrids are often sterile or only set a few viable seeds (1). the reason for this is often the large genetic distance between the species.
Molecular markers provide an excellent means to quantify genetic differences between, but also within, the species. Molecular markers are neutral and independent of the genotype. In this study we used RAPD markers to estimate genetic diversity within and between the speciesCucurbita pepo, C. Maxima and C. Moschata.
Materials and Methods. Plant material: Six C. pepo genotypes, three from each of two Austrian Breeding Companies, Saatzucht Gleisdorf (pep1, pep2, pep3) and VitroPlant (pep4, pep5, pep6), six C. maxima genotypes of different geographic origin (max1USA, max2 China, max3 Japan, max4 Hungary, max5 France, max6 Mongolia) and six C. moschata genotypes (mos1“Nigerian Local” from Nigeria, mos2 “Nicklow’s Delight” from USA, mos3 “Menina” from Portugal, mos4, mos5, and mos6 from Puerto Rico) were selected for this study.
DNA isolation: DNA isolation was carried out using the QIAGEN Dneasy Plant Mini Kit (http://www.qiagen.com, Cat. Nr. 69103). Primers:26 10mer RAPD primers were used supplied by ABgene (http://abgene.com), and ROTH (http://www.Carl-Roth.de). PCR conditions:100 ng of genomoc DNA were used in 25 μl-volume amplification reactions containing 0.3 μM 10mer random primer, 1x reactions bugger, 1.5 mM MgCl2, 200 μM dNTP and 1 Unit Taq polmerase. For amplification we used a TouchDown Thermocycler (Hybaid, http://www.hybaid.co.uk). Temperature program:Initial denaturation of 60 seconds at 94˚C followed by 34 cycles of 60 seconds at 94˚C, 45 seconds at 36˚C, 30 seconds at 72˚C, finished with a final extension step of 5 minutes at 72˚. Fragment separation: Fragments were separated in 1.5% agarose gels stained with ethidium bromide and photographed with a Polaroid camera. Data analysis: Data were recoded to a present-absent scale (1/0) and then subjected to an UPGMA cluster analysis.
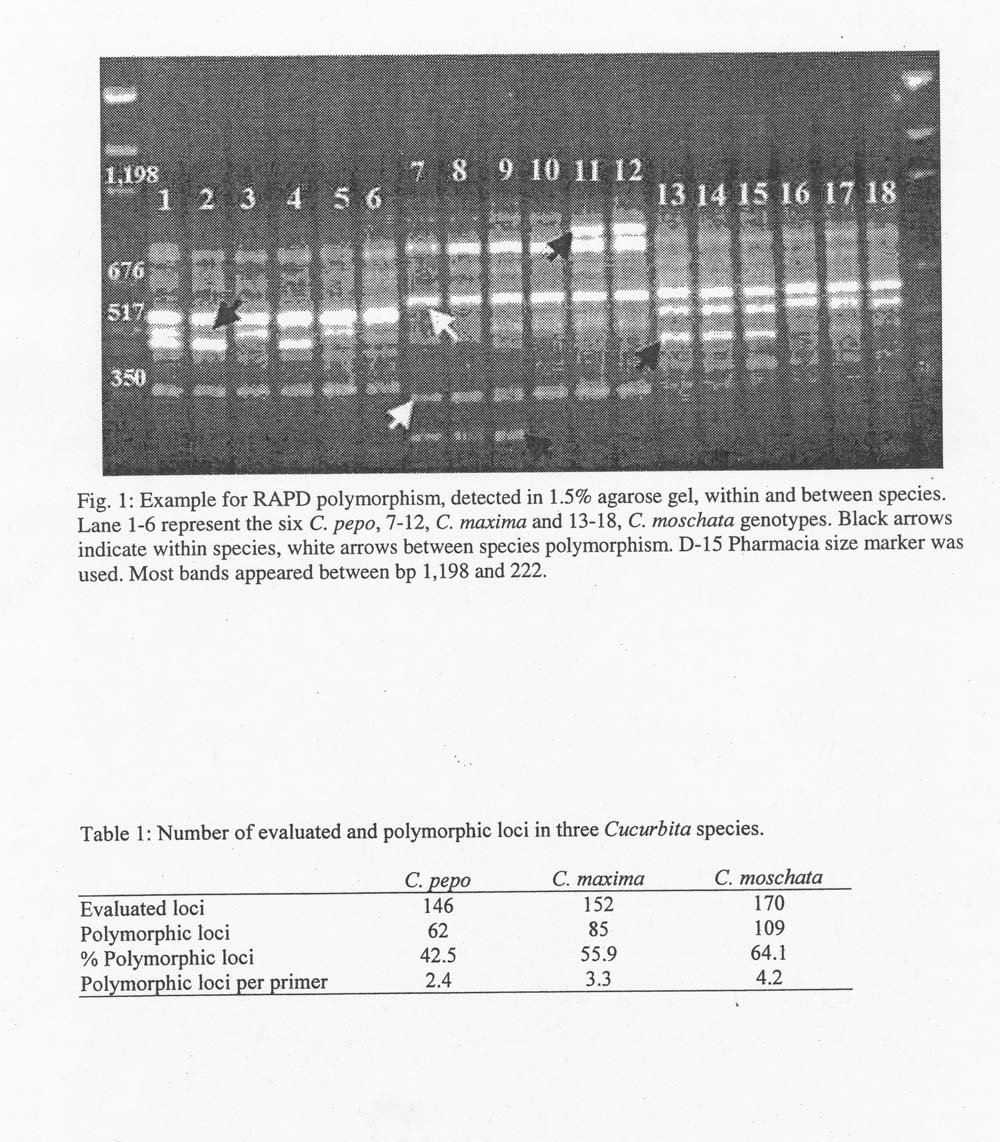
Results and Discussion. RAPDs are dominant markers. Polymorphism is indicated by the presence or absence of a fragment (Fig. 1). Only sharply delineated clear bands were accepted as marker loci. Altogether 379 such loci were recorded. From the total number of loci, ten were monomorphic throughout the three species. Monomorpic loci present in two species, i.e. in C. pepo and C. maxima, in C. pepo and C. moschata, and in C. maxima and C. moschata were 13, 12 and 6 respectively (Fig. 1).Number of evaluated and polymorphic loci in the three species is given in Table 1. Pairwise distances between the 18 genotypes were calculated (Table 2) and a cluster analysis was carried out. (Fig. 2).
The three species show almost equal distance to each other (Table 2). distance within the species is on average less than one fourth of the distance between the species. Comparing the three species clearly the least polymorphic one is C. pepo represented by six Austrian hull-less seeded oil pumpkin genotypes. This is shown by the lowest number of polymorphic loci (Table 1) and by the lowest average distance value (Table 2). This is most probably due to the narrow geographic distribution of the genotypes and a certain drift by selecting for the last hundred years the hull-less seeded types. In an earlier study (5) we detected 3,4 polymorphism per RAPD primer in 20 inbred lines of Styrian oil-pumpkin using 34 primers.
The clearly lower number found in this study (2.4) is probably due to the lower number of genotypes and markers and the fact that the genotypes of the present study were already selected for Austrian growing conditions.
The largest within species distance is exhibited by the six C. moschata genotypes. This group has the highest number of polymorphic loci, the highest number of markers per primer (Table 1), and the highest average distance value (Table 2). This seems to be due to their geographic origin. The three genotypes from Puerto Rico (kindly made available by Dr. Linda Wessel-Beaver) are obviously closely related like the oil pumpkin genotypes from Austria. “Nigerian Local” is a “runaway”, a comparable difference can not be seen in the group of C. maxima with the widest geographic distribution. The large distance of “Nigerian Local” to “Menina” (67 in Table 2) is of special interest, because these genotypes are the two known sources of resistance genes against ZYMV in pumpkin. Because of the large distance between the two genotypes it is possible that these resistance genes are different and therefore suitable for pyramiding in C. pepo.
Table 1. Number of evaluated and polymorphic loci in three Cucurbita species.
| C. pepo | C.maxima | C. moschata | |
| Evaluated loci | 146 | 152 | 170 |
| Polymorphic loci | 62 | 85 | 109 |
| % Polymorphic loci | 42.5 | 55.9 | 64.1 |
| Polymorphic loci per primer | 2.4 | 3.3 | 4.2 |
Table 2. Squared Euclidean dissimilarity between 18 Cucurbita genotypes based on binary data of individual RAPD loci.
| pep1 | pep2 | pep3 | pep4 | pep5 | pep6 | max1 | max2 | max3 | max4 | max5 | max6 | mos1 | mos2 | mos3 | mos4 | mos5 | |
| pep2 | 30 | ||||||||||||||||
| pep3 | 30 | 20 | Average: | ||||||||||||||
| pep4 | 33 | 25 | 33 | pep: 29 | |||||||||||||
| pep5 | 34 | 28 | 30 | 31 | max: 39 | ||||||||||||
| pep6 | 33 | 19 | 33 | 28 | 21 | mos: 48 | |||||||||||
| max1 | 167 | 173 | 173 | 178 | 171 | 162 | pep/mos: 184 | ||||||||||
| max2 | 178 | 184 | 184 | 189 | 182 | 173 | 29 | pep/max: 176 | |||||||||
| max3 | 172 | 178 | 178 | 183 | 174 | 165 | 41 | 32 | mos/max: 182 | ||||||||
| max4 | 171 | 177 | 177 | 182 | 173 | 164 | 46 | 41 | 39 | ||||||||
| max5 | 172 | 178 | 178 | 183 | 174 | 165 | 47 | 36 | 38 | 34 | |||||||
| max6 | 176 | 184 | 182 | 189 | 180 | 171 | 45 | 38 | 40 | 43 | 34 | ||||||
| mos1 | 182 | 186 | 184 | 187 | 188 | 183 | 169 | 172 | 168 | 173 | 172 | 172 | |||||
| mos2 | 183 | 183 | 183 | 188 | 187 | 182 | 182 | 189 | 181 | 188 | 189 | 187 | 67 | ||||
| mos3 | 179 | 177 | 177 | 184 | 179 | 176 | 174 | 183 | 175 | 180 | 179 | 179 | 67 | 36 | |||
| mos4 | 184 | 184 | 182 | 189 | 186 | 183 | 179 | 188 | 180 | 185 | 184 | 186 | 76 | 45 | 41 | ||
| mos5 | 184 | 184 | 184 | 189 | 188 | 185 | 183 | 190 | 184 | 189 | 188 | 192 | 78 | 43 | 39 | 20 | |
| mos6 | 183 | 183 | 181 | 188 | 187 | 182 | 182 | 191 | 183 | 188 | 189 | 189 | 75 | 44 | 38 | 23 | 23 |

Acknowledgement. The stay of the senior author in our department was sponsored by a SOKRATES-ERASMUS stipendium of the EU. His present address is: Mendeleum, Horticultural Faculty in Lednice, Mendel Agricultural Univ., Brno, Czech Republic. We wish to thank Martin Pachner for his excellent technical assistance.
Literature Cited
- Baggett, J.R., 1979. Attempts to cross Cucurbita moschata (Duch.) Poir. ‘Butternut’ and C. pepo L. ‘Delicata’. Cucurbit Genetics Coop. Rpt. 2:32-34.
- Gilbert-Albertini, F., H. Lecoq, M. Pitrat, and J.L. Nicolet, 1993. Resistance of Cucurbita moschata to watermelon mosaic virus type 2 and its genetic relation to resistance to zucchini yellow mosaic virus. Euphytica 69:231-237.
- Munger, H.M., and R. Provvidenti, 1987: Inheritance of resistance to zucchini yellow mosaic virus in Cucurbita moschata. Cucurbit Genetics Coop. Rpt. 10:80-81.
- Paris, H.S., S. Cohen, Y. Burger, and R. Yoseph, 1988: Single-gene resistance to zucchini yellow mosaic virus in Cucurbita moschata. Euphytica 37:27-29.
- Stachel, M., Gy. Csanadi, J. Vollmann, and T. Lelley, 1998: Genetic diversity in pumpkins (Cucurbita pepo L.) as revealed in inbred lines using RAPD markers. Cucurbit Genetics Coop. Rpt. 21:48-50.
- Whitaker, T.W., and R.W. Robinson, 1986.: Squash breeding. In: Breeding vegetable crops. Ed.: M.J. Bassett. Avi, Westport, Ct.