Cucurbit Genetics Cooperative Report 23:89-95 (article 31) 2000
J. Brent Loy
Department of Plant Biology, University of New Hampshire, Durham, NH 0324, U.S.A.; [email protected]
Abstract: Seeds of Cucurbita pepo L. are high in protein and oil, and a good source of potassium, phosphorous, some of the minor elements, and some of the B vitamins. The seeds are also large and abundant in fruits. Nonetheless, they are underutilized as a food crop, probably owing to the thick, leathery seed coat (hull) that cannot be removed easily be mechanical means. The existence in this species of a mutant seed phenotype characterized by an unlignified, thin seed coat has provided a genetic means by which breeders can develop new germplasm with essentially hull-less seeds that can be more easily exploited as a food and source of vegetable oil. To better exploit hull-less seeded cultivars as a food crop, we have been conducting research on seed development in this species. We subdivide seed development into three phases: (1) expansion and biomass accumulation in seed coats and differentiation of the embryo; (2) rapid embryo enlargement and depletion of seed coat reserves; and (3) Biomass accumulation (seed fill) in embryos. Biomass accumulation in seed coat peaks at 20 days post-anthesis (PA), coinciding with maximal fruit expansion. In both hulled and hull-less seeds extensive amounts of nonstructural constituents (starch, lipids, sugars, amino acids) accumulate in seed coats between 0 to 20 days PA. In normal seeds, secondary cell wall thickening begins early in seed coat development and is characterized by extensive lignification and cell wall thickening in hypodermal, sclerenchymatous and parenchymatous tissues by 20days PA. Secondary wall thickening continues as seeds mature. In hull-less genotypes there us a reduction in lignification of tissues, the difference being first detectible by lignin staining at 10 days PA, but becoming increasingly apparent during seed development. Cellulose accumulation is also reduced in hull-less as compared to hulled genotypes early in seed development, but unlike lignin, this difference is not accentuated in mature seeds. Rapid embryo enlargement occurs between 20 to 40 days PA. Seed fill increases more or less linearly between 25 to 60 days PA; however, rates of seed fill and the duration of seed fill may vary according to environmental conditions during fruit growth and seed development. In fruits harvested prematurely, assimilates stored in mesocarp tissue can be remobilized to the developing seed. Changes in color of the fruit exocarp normally associated with maturation of pumpkins do not necessarily coincide with the time of maximum seed fill.
Introduction: Seeds are a rich source of nutrients for human consumption, and as such, serve as the main food base for most of the world population. Seeds within the genus Cucurbita contain 32 to 37% protein and 42 to 50% oils (6), and also contain relatively high amounts of potassium, phosphorous and zinc (9) and significant quantities of niacin and thiamine (7). Thus, they represent a highly nutritious and potentially important seed crop for human use. However, because of the thick, leathery seed coat associated with most cultigens, and the difficulty in decorticating the seed, Cucurbita seeds have been underutilized as a human food. A recessive mutation (n) in C. pepo, in which secondary cell wall constituents are much reduced within the outer seed coat tissues (5, 14), provides a convenient genetic trait for developing hull-less seeded cultigens, the seeds of which can be more readily utilized as a human food source. These hull-less or ‘naked seeded’ cultigens of pumpkin may have been propagated for over 100 years in Austria (H. Teppner, personal communication) but the first published reference to genetically hull-less or ‘schalenlosen’ seed in C. pepo of which I am aware was in 1934 by an Austrian scientist, Tschermak-Seysenegg (16). Because pumpkin seed oil was highly valued in austria and other parts of Eastern Europe for use in salad dressings, the utility of hull-less seeded strains for more efficient extraction of the oil was immediately recognized.
Seed coat composition and function in hulled and hull-less cultigens of C. pepo: Morphologically, seed coats of cucurbita pepo are subdivided into two layers that can be easily separated in mature seed, a relatively thick, leathery outer layer and a thin, membranous-like, green inner layer. In 1909 Barber (3) reviewed the major nineteenth century contributions to cucurbit seed morphology, and provides an accurate illustration of the major seed coat tissues in Cucurbita pepo. Singh and Dathan (11) give a more detailed account of the derivation of the five zones or tissue types comprising the seed coat layers. The inner seed coat consists of several layers of thin-walled, relatively large, chlorenchyma cells. The outer seed coat consists of tangentially elongated, thin-walled, nonlignificed epidermal cells, 3 to 5 layers of small, extensively lignified hypodermal cells, a single layer of large, heavily lignified schlerenchymatous cells, and one or two layers of small, partially lignified parenchymatous and aerenchymatous cells. These tissue layers are formed by 10 days post-anthesis (PA), but the palisade epidermal cells are not fully elongated until 15 to 20 days PA (12). In mature desiccated seeds, the inner layer of chlorenchyma tissue compresses into a thin, membranous-like chlorophyllous layer (14). This layer usually remains intact when seeds are decorticated by hand. In the outer seed coat of mature, dried seed, the epidermis largely collapses, but the lignified hypodermal, sclerenchymatous, and arenchymatous tissues maintain their integrity and form the hard,m leathery hull characteristic of cucurbita seed.
Heinisch and Ruthenberg (5) made a detailed anatomical study of the seed coat in hulled and hull-less genotypes of pumpkin and reported that in ‘naked seed’ genotypes all seed coat layers were present, but thickening and lignification of cell walls were reduced,. They as well as other early investigators (8, 19, 19) noted genotypes in segregating populations with different degrees of cell wall thickening.
Stuart and Loy have conducted more detailed anatomical studies and have also analyzed seed coat composition in hulled and hull-less cultigens. At 10 days post-anthesis, prior to appreciable secondary wall development, hulled and hull-less genotypes are nearly indistinguishable (14), with the exception of slight staining with phloroglucinal (indicative of lignin) in normal genotypes. However, by 15 days post-anthesis, phlorogluycinal-positive staining within hypodermal, sclerenchymatous and aerenchymatous tissue is clearly much greater in hulled as compared to hull-less genotypes (12). The disparity in secondary cell wall thickening between hulled and hull-less genotypes continues to magnify as the seeds mature.
Quantitative estimates of the structural (cell wall) constituents of seed coats at 20 days PA and in mature seeds are given in Table 1. Henicelluloses, pectins and cellulose are the predominant cell wall constituents in normal seed coats at 20 days PA. Mutant seed coats exhibit normal amounts of pectins and hemicelluloses, but have much reduced amounts of cellulose and, especially, lignin as compared to hulled genotypes. Pectins and to a lesser extent hemicelluloses are largely degraded between 20 days PA and seed maturity. The reduction in hemicelluloses is much greater in hull-less as compared to hulled seeds. This may be because there is greater accessibility within secondary cell walls of hull-less than of hulled genotypes to hydrolytic enzymes that degrade cell wall polysaccharides (see 1, 2). In normal, hulled seed there is about a 3-fold increase in lignin and in some cases a slight increase in cellulose accumulation between 20 days PA and seed maturity. The most pronounced differences between seed coat genotypes at seed maturity are 60 to 70% reductions in cellulose and 79 to 88% reductions in lignin content in hull-less as compared to hulled seed coats (Table 1). The reduction in cellulose in hull-less as compared to normal seeds at maturity may well be at least partly due to greater degradation by hydrolytic enzymes as suggested for the hemicelluloses. It should be noted that the hull-less cultigens used in Stuart’s studies (12, 13) exhibited some cell wall development in the outer seed coat, especially along the seed margins. This is in contrast to most of the Styrian cultigens I have observed, and to several of our own breeding lines, in which lignified tissues of the outer seed coat are virtually absent.
Nonstructural constituents, starch, lipids, sugars, free amino acids, and soluble proteins, are in great abundance in seed coats at 20 days PA (Table 2). Starch is especially abundant in chlorenchymatous tissues in both hulled and hull-less genotypes, but is also relatively abundant in hypodermal and parenchymatous cells of hull-less strains (14, 17). It is conceivable that in hulled as compared to hull-less genotypes, the starch in these latter two tissues may be more rapidly utilized for secondary wall formation. Between 20 days PA and seed maturity the nonstructural constituents are largely depleted. In hulled seeds it is presumed that some of these assimilates are utilized for lignin synthesis. In both genotypes, however, these constituents appear to serve as a major reservoir of assimilates for the developing embryo.
Stages of seed development:
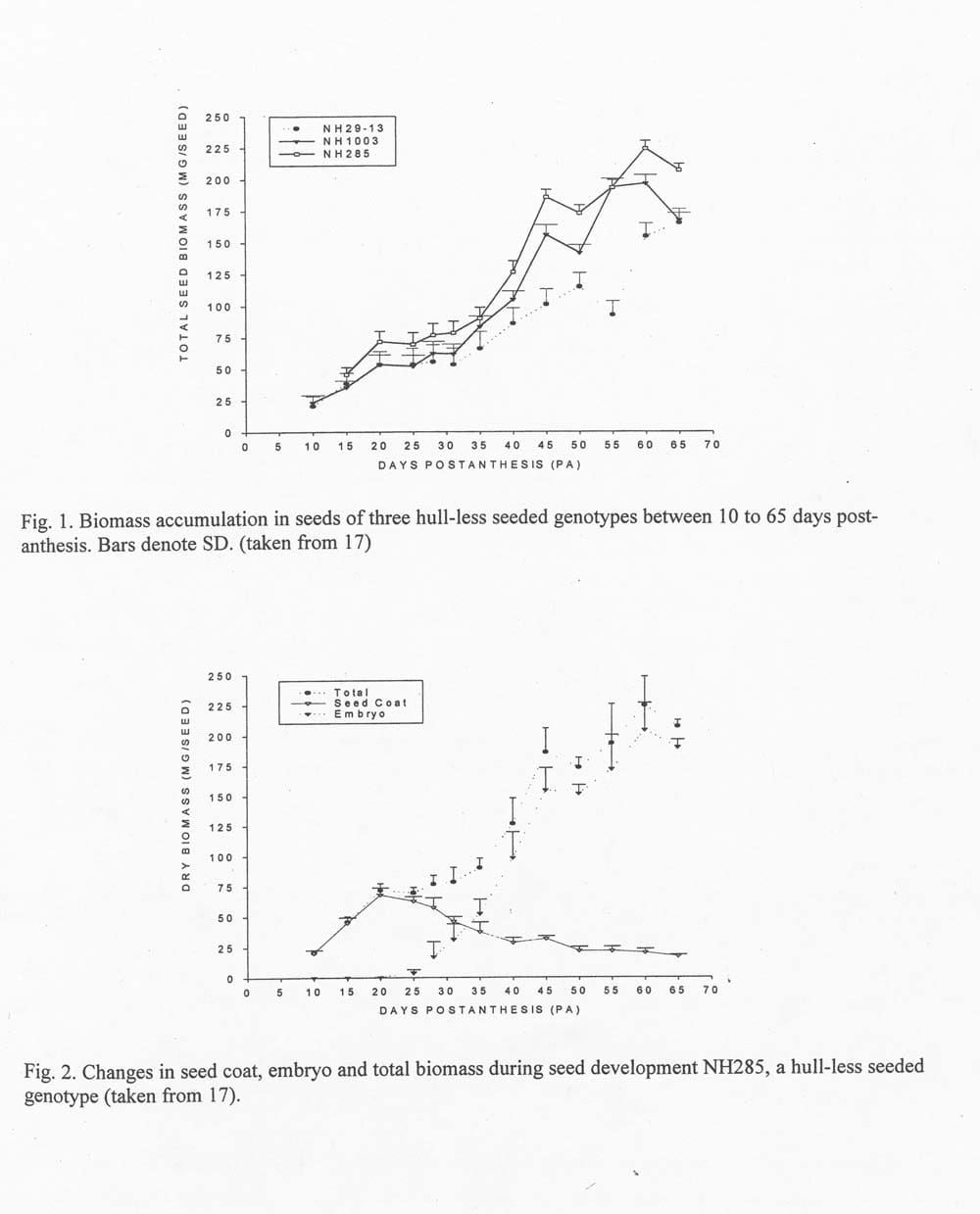
Seeds exhibit a relatively linear increase in total biomass from 10 days post-anthesis (PA) until seed maturity at 55 to 60 days PA (Fig. 1.) However, different organs within the seed display different periods of development and rates in biomass accumulation (18). As such, seed development in C. pepo can be conveniently subdivided into 3 distinct, but overlapping stages: (1) 0 to 20 days PA – period of expansion and accumulation of reserves in seed coats, endosperm development and differentiation of embryos: (2) 20 to 40 days PA – period of rapid embryo enlargement, reduction in endosperm fresh weight and decrease in seed coat nutrient reserves; and (3) 25 to 60 days PA – period of accumulation of seed storage reserves.
Fresh weight and biomass of seed coats increases rapidly after fertilization and peaks at 20 days PA, after which the decrease slows until seeds reach maturity. Embryos remain relatively small and difficult to detect with the naked eye until about 20 days PA. Shortly thereafter, embryos enlarge rapidly and fill in the seed coat cavity between 35 to 40 days PA. The period of rapid cotyledon enlargement may vary considerably even within a single fruit, but according to our observations, rarely begins until after 20 days PA and cytologically appears complete by or before 40 days PA, but occasionally 3 mm long embryos were recovered at that stage (17). Quantitatively, embryos show nearly linear increases in fresh weight until about 45 days PA, after which growth usually slows until seed maturity at 60 days PA. Later stages of embryo expansion are often coincident with collapse of perisperm and inner seed coat tissues. We have on occasion, however, observed some fresh seeds at 60 days PA in which the perisperm and inner seed coat tissues are still largely intact, because of poor seed fill.
The patterns of biomass increase among seed organs can be expected to vary with differences in growing conditions. In our 1997 experiment seed biomass at 35 days PA was 40 to 42% of that in mature seed among three genotypes. But in a test of 11 experimental, hull-less seeded hybrids in 1999, seed biomass at 35 days PA averaged 55% of that of mature seed. The 1999 growing season was unusually warm and seed development occurred between about 15 July to 15 September. In 1997 fruit and seed development occurred between 28 July and 30 September under much cooler but fairly normal conditions for our region.
Fruit maturity and biomass changes in seeds from stored fruit: During crop growth plants ideally remain healthy and canopy photosynthesis remains active until fruit and seeds mature. Our observations on culture of pumpkins over the past 20 years suggest that it is quite common for plants to senesce prematurely before fruits are deemed mature. Relative to this phenomenon, we have been confronted with two problems when dealing with our culture and evaluation of hull-less seeded hybrids: (1) from visual observations of rind color, can we tell when fruit and seeds within the fruit are mature, and (2) if vines senesce prematurely, will fruit and seeds continue to develop and mature, and if so, to what extent.
The first problem was relatively easy to tackle, but resulted in a few surprises. We compared two inbred lines, NH285 and NH29-13, and their F1 hybrid. We presumed from previous field observations that NH285 was late maturing. In contrast, we assumed that NJ29-13 matured early because of early changes in fruit color from green to dark orange. In our comparative studies in 1997, the skin or rind of NH29013 fruit indeed showed extensive changes from green to almost full orange coloration between 30 to 40 days PA (17, 18). On the other hand, NH285 fruit exhibited only subtle changes in skin color from the initial pattern of alternating light and dark green stripes. AT 45 days PA about half of the fruit exhibited some tinges of orange color light green portion of fruit, and after 60 days most fruit were light orange and green striped on the surfaces exposed to the sun. NH1003, the F1 hybrid, were striped, but the striping was less distinct than that of NH285. By 40 days PA some fruits exhibited light orange and green stripes, and by 55 days PA most fruits were yellow-orange with no prominent striping. The different patterns of fruit coloration cited above were often not correlated with seed maturation. In all three cultigens embryo biomass increased significantly between 50 to 60 days PA, at a time when all fruits of NH29-13 and most fruits of NH1003 appeared mature. The changes in embryo biomass during this late period of maturation were astonishing: 43% increase in NH29-13, 47% increase in NH1003, and a 35% increase in NH285, the presumably late maturing cultigen.
Fruit mesocarp tissues reach peak biomass in C. pepo at about 30 days PA (4,18), at a time when seed fill is just beginning to accelerate. Thus, we hypothesized that fruit mesocarp reserves might serve as a source of assimilates for developing seeds under conditions in which leaf canopy production of photosynthates is limited. To test this, we excised fruit prematurely (35 to 50 days PA) from field-grown plants and stored them in a glasshouse for 10-day periods; changes in seed fill were compared between stored and intact fruits (18). In two sampling periods, 35 to 45 days PA and 40 to50 days PA, seed fill in stored fruits of NH29-13 was 12,7 and 20.5% lower, respectively, than that of intact fruit. In three sampling periods (35-45, 40-50, and 45-55 days PA) of NH1003, seed fill in stored fruit was 17.2, 16.6 and 58.3% lower than that in corresponding intact fruit. In two sampling periods (40-50, 45-55) for NH285, seed fill in stored fruit was essentially the same as in intact fruit. In more recent studies completed in 1999, we compared seed fill in intact fruit to those excised and stored in the field for at least 25 days, from 35 to 60-65 days PA. In this study, comparing 11 hybrids, seed fill in stored fruits averaged 20% less than that of intact fruit, even though seed biomass was 55% complete by 35 days PA. In 9 of the 11 hybrids for which we had reliable data, the increased seed fill (biomass) in stored fruits accounted for between 32 to 54% of the estimated loss in mesocarp biomass during the storage period. The results thus show that in absence of a supply of photosynthates to developing seeds during the seed fill stage, assimilates in fruit tissues can be remobilized to the seed.
Conclusions: The dynamics of fruit and seed development in cucurbita pepo pumpkins are not only important from a scientific viewpoint, but also for developing optimum cultural systems for maximizing and sustaining fruit and seed production. Fruit reach optimum size at about 20 days PA, and this also coincides with the period of peak fresh weight and biomass of seed coats (18). The function of seed coats is much broader than just a protective covering for seed embryos. All nutrients transported from the fruit tissues to the developing embryo enter the seed via the vasculature connecting the placental tissue to the micropylar end of the seed and running along the margins of the seed coat. Subsequent transport to the embryo must occur apoplastically and/or symplastically through the seed coat and perisperm tissues, and apoplasticaly from the perisperm to the endosperm enclosing the embryo. Starch, lipids, pectins, sugars and amino acids are abundant in seed coats during early seed development, so seed coats appear to serve as a major reservoir of nutrients for later embryo expansion and seed fill. It is also sometimes not appreciated that in C. pepo and other cucurbits seed size is largely maternally regulated or determined. Maximum seed coat size at 20 days PA largely delimits the degree of embryo expansion and final seed size. This phenomenon is true for both hull-less and hulled genotypes. Compared to normal hulled seed, the seed coat in hull-less genotypes does not offer a strong physical barrier to embryo expansion, so differences in water potentials between embryo tissue and adjacent maternal tissues (perisperm and inner seed coat) likely regulate final embryo size in developing seed.
Because seed coat the fruit expansion occur simultaneously, the degree of locule expansion influences seed coat expansion and final seed width and length dimensions. For example, in small fruit (0.5 to 1.2 kg) we rarely obtain seeds larger than 200 mg; whereas in all of our large-seeded lines, fruit size is greater than 2.0 kg. Nonetheless, breeding liens with small fruit but endowed with genes for large seed, will produce large seeds under conditions of low seed set.
A considerable portion of final seed biomass may accumulate during the last 10 days of seed development and maturation which, in some cases, occurs much later than when fruit appear mature according to changes in skin color. Therefore, determination of the extent of maximum seed fill is critical for maximizing seed yields in commercial production. furthermore, because nutrient reserves in fruit tissues reach a maximum at 30 days PA and can be remobilized to the developing seed, acceptable seed yields may be obtained in production fields under conditions in which plants senesce prematurely due to disease or other stress conditions.
Table 1. Structural (cell wall) constituents of seed coats in 20-day post-anthesis and mature seeds of normal and hull-less cultivars of pumpkin.z
| 20-day PA seed coats | Mature seed coats | |||
| Hulled | Hull-less | Hulled | Hull-less | |
| Constituent | mg per seed coat | |||
| Proteiny | 5.07 | 4.24 | 6.51 | 2.94 |
| Pectins | 9.65 | 8.45 | 0.85 | 0.55 |
| Hemicellulosesx | 10.15 | 10.25 | 4.94 | 1.63 |
| Cellulose | 8.65 | 3.70 | 9.65 | 2.99 |
| Lignin | 3.20 | 0.60 | 8.80 | 1.37 |
z Values extrapolated from data of Stuart and Loy (13, 14) and based on averages from analysis of two hulled cultivars (Small Sugar and Jack o’ lantern) and two hull-less cultivars (Tricky Jack and 293A).
y Proteins from mature seed estimated from N content of digested samples x 6.25, and considered to be largely those associated with the cell wall. Proteins from 20-day PA seeds were fractionated into soluble and bound fractions, with only the later given in Table 1.
x Values represent combined fractions of hemicelluloses, HcA and HcB.
Table 2. Nonstructural constituents in seed coats of 20-day post anthesis (PA) and mature seed of hulled and hull-less cultivars of pumpkin.z
| 20-day PA seed coats | Mature seed coats | |||
| Hulled | Hull-less | Hulled | Hull-less | |
| Constituent | mg per seed coat | |||
| Soluble protein | 2.25 | 2.19 | – | – |
| Free amino acids | 3.07 | 3.43 | 0.17 | 0.15 |
| Reducing sugars | 5.20 | 4.53 | 0.24 | 0.25 |
| Phenolics | 0.24 | 0.27 | 0.03 | 0.03 |
| Starch | 9.55 | 10.82 | 0.97 | 0.22 |
| Lipids | 6.65 | 5.11 | 0.93 | 1.21 |
z Values from data of Stuart (12), and based on averages from analysis of two hulled cultivars (Small Sugar and Jack o’ lantern) and two hull-less cultivars (Tricky Jack and 293A).
Literature Cited
- Bailey, R.W. and D.I.H. Jones. 1971. Pasture quality and ruminant nutrition. III. Hydrolysis of rye grass structural carbohydrates with carbohydrases in relation to rumen digestion. N.Z. J. Agr. Res. 14: 847-857.
- Bailey, R.W. and S.E. Pickmere. 1975. Alkali solubility of hemicelluloses in relation to delignification. Phytochemistry 14:501-504.
- Barber, K.G. 1909. Comparative histology of fruits and seeds of certain species of Cucurbitaceae. Bot. Gaxz. 47:263-310.
- Culpepper, C.W. and H.H. Moon. 1945. Differences in the composition of the fruits of Cucurbita varieties at different ages in relation to culinary use. J. Agr. Res. 71:111-136.
- Heinisch, O. and M. Ruthenberg..1950. Die Bedeutung der samenschale fur die Zuchtung des Okurbis. Z. Pflanzen. 29:159-174.
- Jacks, T.J., T.P. Hensarling and L.Y. Yatsu. 1972. Cucurbit seeds: I. Characterizations and uses of oils and proteins. A review. Econ. Bot. 26:135-141.
- Mansour, E.H., E. Dworschak, A. Lugase, E. Barna and A. Gerely, 1993. The nutritive value of pumpkin (Cucurbita pepo Kakai 35) seed products. J. Sci. Food Agr. 61:73-78.
- Mudra, A. and D. Neumann, 1952. Probleme und Ergebnisse der Munchenberger Olkurbiszuchtung Zuchter 22:99-105.
- Robinson, R.G. 1975 Amino acid composition of sunflower and pumpkin seeds. Agron. J. 67:541-544.
- Schoeniger, G. 1950. Genetische Untersuchungen und Cucurbita pepo. Zucher 20:321-336.
- Singh, D. and A.S.R. Dathan. 1972. Structure and development of seed coat in cucurbitaceae. VI. Seeds of Cucurbita. Phytomorphology 22:29-45.
- Stuart, S.G. 1981. Comparative studies of testa development in normal and hull-less seed strains of Cucurbita pepo L. M.S. Thesis, University of New Hampshire, Durham, N.H.
- Stuart, S.G. 1983. Comparative biochemical and genetic studies of testa development in normal and hull-less phenotypes of pumpkin (Cucurbita pepo L ). Ph.D. thesis, University of New Hampshire, Durham, NH, U.S.A.
- Stuart, S.G. and J.B. Loy. 1983. Comparison of testa development in normal and hull-less seeded strains of cucurbita pepo L. Bot. Gaz. 144:491-500.
- Stuart, S.G. and J.B. Loy. 1988. Changes in testa composition during seed development in cucurbita pepo L. Plant Physiol. (Life Sci. Adv.) 7:191-195.
- Techermak-Seysenegg, E. 1934. Der Kurbis mit schalenlosen samen, eine beachtenswerte Olfruct. Wiener Landwirts. Zeit. 84:7-15.
- Vining, K.V. 1999. Seed development in hull-less seeded pumpkin (Cucurbita pepo L.), M.S. Thesis, University of New Hampshire, Durham, NH, U.S.A.
- Vining, K.V. and J.B. Loy. 1998. Seed development and seed fill in hull-less seeded cultigens of pumpkin (Cucurbita pepo L.), p. 64-69. In: J.D. McCreight (ed.), Cucurbitaceae 98: Evaluation and enhancement of cucurbit germplasm. ASHS Press.
- Weilung, F. and E. Prym von Becherer, 1950. Zur Factorenanalyse der Testaausbeldung beim Kurbis. Ber. Deut. Bot. Ges. 63:147-148.