Cucurbit Genetics Cooperative Report 6:5-7 (article 2) 1983
M. D. Edwards and R. L. Lower
University of Wisconsin, Madison, WI 53706
Previous reports have described seed quality problems associated with the compact plant type (cp cp) in cucumbers (1). Variability exists in compact populations for emergence potential of seeds as well as for associated traits (2). Emergence percentage in a reference compact population has been shown to exhibit a moderate level of heritability and to be under the control of the genotype of the seed parent, i.e. maternally controlled (3).
Inbred compact lines have been obtained by recurrent backcrossing of a compact donor into each of five commercial pickling inbreds. Under most conditions, the quality of seeds produced on these inbred compact plants has been poorer than that of a reference compact population. The reason for this particularly poor seed quality is unknown.
Cucumbers have been reported to exhibit measurable inbreeding depression for some traits (4, 5, 6). The magnitude of inbreeding depression observed has usually not been great. Consequently, the crop is not generally considered to suffer from inbreeding depression. This study was conducted to determine if the extremely low seed vigor observed for inbred lines of compact cucumbers might be attributable to inbreeding depression.
F1 and F2 seed lots were produced in the greenhouse for each of seven crosses among five inbred lines of compact cucumbers. Because of the poor quality of seed produced on cp cp plants, all seed lots were produced on Cp cp plants which segregated cp cp individuals among the progeny. Parental, F1 and F2 generations were sown in the field in ten-plant plots on June 16, 1982 at Hancock, WI. Spacing was 30 cm between hills and 1.5 meters between rows. Plots were planted in a randomized complete block design with three blocks. Hills were over-seeded and each was thinned to one compact plant at the two-leaf stage. Because of the maternal regulation of seed quality (3), plants were allowed to open-pollinate to produce fruit.
At maturity, seed from one fruit was harvested from each plant. Seed from all fruit in each plot were pooled to form a seed bulk. Seed bulks were fermented two days at room temperature then washed, dried and packaged. After 15 months of storage in laboratory conditions, two 100-seed samples were counted from each bulk, weighed, treated with a fungicide and sown 2.5 cm deep in a heated sand bench. Emergence testing was conducted using a randomized complete block design with two blocks. All three field bulks for each treatment were represented in each block. Emergence percentage and mean number of days to emergence were recorded for each sample.
Mean values of dependent variables for each generation and cross are presented in Table 1. The F1 and F2 generation values for seed weight exceeded the mid-parent value in all seven crosses. F1 values also exceeded the high parent for four of the seven crosses. The consistent relationship between the inbreeding coefficient and seed weight can be seen in Figure 1. Lines represent the least-squares regression relationships for each cross, where mid-parent values are used for the parental generation.
Table 1. Mean values of seed traits for seedlots produced on various maternal parents
Maternal Source |
Cross Designation |
Seed Weight
|
Emergence
|
Mean # Days
|
|---|---|---|---|---|
| Inbreds | ||||
| Addis cpcp | .39 | 1 | 7.2 | |
| GY2 cpcp | .83 | 60 | 5.8 | |
| Chipper cpcp | .70 | 7 | 7.2 | |
| GY14 cpcp | .70 | 20 | 6.1 | |
| 8L cpcp | .63 | 9 | 6.7 | |
| F1’s | ||||
| Addis x Chipper | A | .78 | 9 | 6.8 |
| GY2 x Addis | B | .66 | 17 | 6.4 |
| GY2 x Chipper | C | .78 | 22 | 6.6 |
| GY2 x 8L | D | .90 | 47 | 5.9 |
| GY14 x Addis | E | .67 | 17 | 6.6 |
| GY14 x GY2 | F | .89 | 57 | 5.3 |
| GY14 x 8L | G | .78 | 17 | 7.2 |
| F2’s | ||||
| Addis x Chipper | A | .71 | 25 | 6.4 |
| GY2 x Addis | B | .60 | 6 | 6.9 |
| GY2 x Chipper | C | .66 | 25 | 6.4 |
| GY2 x 8L | D | .79 | 48 | 6.0 |
| GY14 x Addis | E | .66 | 3 | 7.5 |
| GY14 x GY2 | F | .79 | 49 | 5.3 |
| GY14 x 8L | G | .69 | 17 | 6.0 |
| LSD .05 | 19 |
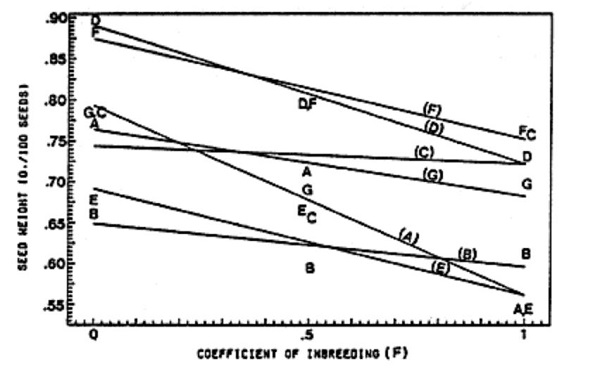
Figure 1. Effect of inbreeding on level of seed weight.
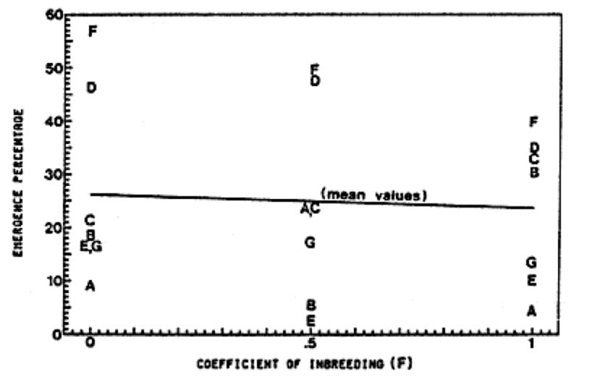
Figure 2. Effect of level of inbreeding on emergence percentage
The relationship between inbreeding and emergence percentage is less clear (Figure 2). Although the slope of the least-squares regression line for generation means across the seven crosses is negative, specific crosses do not show consistently decreased emergence percentage with increased levels of inbreeding.
These data clearly indicate inbreeding depression effects for seed weight in compact cucumbers. Whether inbreeding also affects emergence percentage is less evident. The coefficient of variation (CV) for emergence percentage is much larger than that for seed weight, 54 and 11 percent, respectively. The large error variance for emergence percentage makes it difficult to accurately describe trends associated with inbreeding. Previous studies have documented a strong phenotypic association between seed weight and emergence percentage within a heterogeneous compact population (2). It seems likely that with greater experimental precision, inbreeding would be found to be associated with poorer emergence percentages.
However, the average emergence percentage of seed produced on F1 hybrids in this study is considerably poorer than values obtained for seed lots from open-pollinated compact plants in an adjacent plot. This suggests that factors other than inbreeding are probably also involved in the poor emergence performance of inbreds utilized in this investigation.
Literature Cited
- Edwards, M. D. and R. L. Lower. 1981. Investigations into the characteristics of seeds from compact cucumber plants. Cucurbit Genetics Coop. Rpt. 4:2–4.
- Edwards, M. D. and R. L. Lower. 1981. Variability for seed quality among fruit from individual compact cucumber plants. Cucurbit Genetics Coop. Rpt. 4:4–5.
- Edwards, M. D. and R. L. Lower. 1982. The genetic regulation of several seed traits in compact (cpcp) cucumbers—maternal vs. embryonic control. Cucurbit Genetics Coop. Rpt. 5:8–9.
- El-Shawaf, I. I. S. and L. R. Baker. 1981. Inheritance of parthenocarpic yield in gynoecious pickling cucumber for once-over mechanical harvest by diallel analysis of six gynoecious lines. J. Amer. Soc. Hort. Sci. 106:359–364.
- Ghaderi, A. and R. L. Lower. 1979. Heterosis and inbreeding depression for yield in populations derived from six crosses of cucumber. J. Amer. Soc. Hort. Sci. 104:564–567.
- Hutchins, A. E. 1938. Some examples of heterosis in cucumber, Cucumis sativus L. Proc. Amer. Soc. Hort. Sci. 36:660–664.