Cucurbit Genetics Cooperative Report 18:7-9 (article 4) 1995
J.E. Staub, Jeff Bacher and Linda Crubaugh
Vegetable Crops Research, USDA/ARS, Departments of Horticulture, University of Wisconsin-Madison, WI 53706 USA
Introduction. To respond to the need for cultivars suitable for mechanical harvesting our breeding project is manipulating cucumber plant architecture to develop high yielding genotypes. Standard cucumber varieties (gynoecious x monoecious or gynoecious x gynoecious hybrids) possess an indeterminate plant habit (DeDe) and few lateral branches (~ 1 to 2). We are developing all female genotypes which are short in stature (determinate: de) and possess a multiple lateral branching habit (~ 3to 5). This plant type can be sown at relatively high densities (compared to standard indeterminate types and will allow for early, concentrated fruit set on more lateral branches (Staub et al., 1992)
There are two problems inherent to such a breeding project. First, vegetative propagation of determinate types is extremely difficult. Tissue culture has proven ineffective because of recurrent problems with third contamination of selections; rooting of cuttings has not succeeded in producing flowering plants because juvenility can not be restored in determinate plants; and senescing determinate plants rooted by stem layering techniques do not survive transplantation. Controlled self-pollination of all selected plants in a field is impractical early in the season and late season chemically induced sex conversion of mature gynoecious determinate plants to a monoecious flowering habit is impossible.
The second problem is the potential misclassification of mature plants (indeterminate versus determinate types) because of difficulty in identifying the determinate character in a multilateral background. Differences in vine length are mitigated by the removal of apical dominance resulting in plants which are difficult to distinguish. Although de conditions termination of growing points at the terminal whorl, the length of lateral branches of determinate plants is quantitatively inherited and can vary dramatically. Size variation observed in segregating progeny derived from determinate x indeterminate crosses is presented here to demonstrate the difficulty in selecting multiple lateral determinate phenotypes.
Materials and Methods. The determinate unilateral line G421 (R.L. Lower, University of Wisconsin) was mated with the indeterminate multiple lateral line H-19 (University of Arkansas) to produce F2 , F3 , BC1 S2 and BC1 S2 progeny. F3 progeny were produced from random F2 ‘s chosen from the middle of the progeny distribution, and G421 was used as the recurrent parent during backcrossing. Measurements of the mainstem of F2 and F3 greenhouse grown plants were taken about six weeks after transplanting (two-leaf stage). Mainstem measurements of field grown (Hancock, WI) plants sown on 1.5 m row centers with a within row spacing of 0.76 m were taken approximately eight weeks after sowing. F3 plants classified as determinate and indeterminate were self-pollinated to produce F4 lines which were measured in a greenhouse. F4 and BC1 S2 plants were classified for determinate character during the same period.
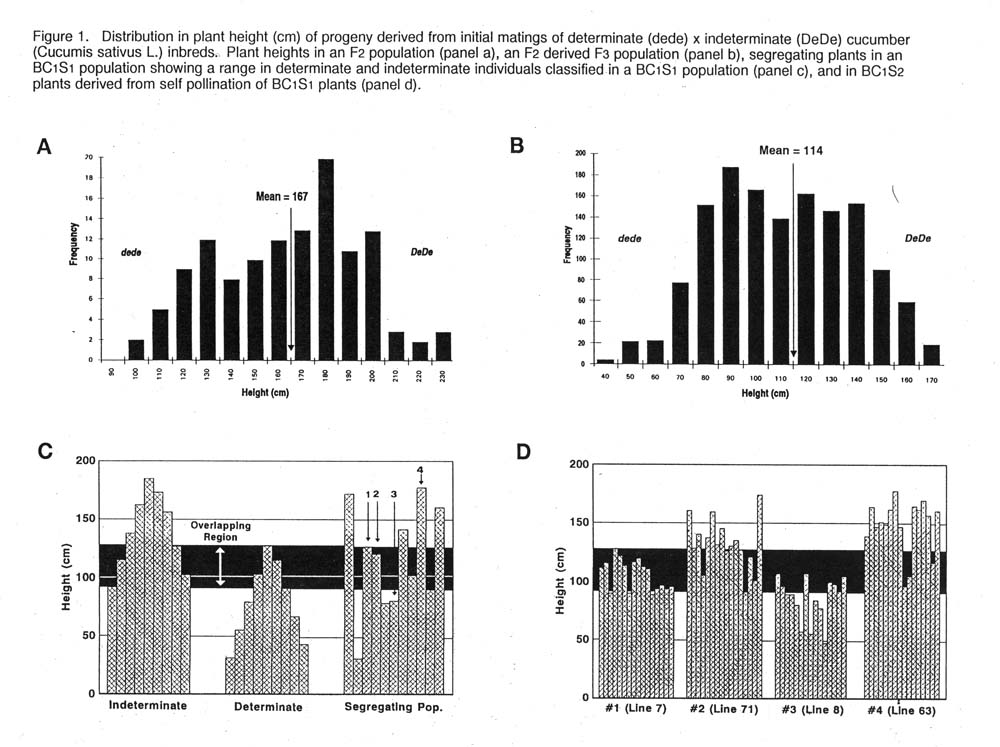
Results and discussion. Progeny segregation in the F2 generation formed a continuous normal distribution (Figure 1, panel a). Mainstem length varied from ~ 100 to 230 cm and having a mean length of 167 cm. Determinate phenotypes were classified as being between ~ 100 to 140 cm and having a mean length of 167 cm. Determinate phenotypes were classified as being between ~ 100 to 140 cm and indeterminates were classified as measuring between ~ 200 to 230 cm. The length of the mainstem was normally distributed in the F3 generation ranging between ~40 to 170 cm and indeterminate were classified as measuring ~ 140 to 170. Plants in the middle of all distributions, regardless of the generation, could not be classified with precision. The growing environment can greatly affect the ultimate length of the mainstem.
The distribution of indeterminate and determinate F4 plants is presented in Figure 1, panel c (far left). A region (“overlapping region”) where plants could not be classified was characterized. Segregating BC1 S2 progeny varied in mainstem length [Figure 1, panel c (far right)] and four were self-pollinated. Measurements of BC1 S2 progeny indicate that BC1 S2 selections made in the overlap region (Lines 7 & 71) were difficult to classified in BC1 S2. In contrast, BC1 S2 progeny resulting from BC1 S2 lines selected beyond the overlapping region (Lines 8 & 63) were comparatively easier to classify. Nevertheless, in each case the mainstem length of some BC1 S2 progeny fell into the overlapping region indicating that stabilization of moderately large determinate lines (~ 90 cm mainstem length) will require judicial selection in advanced generations.
Figure 1. Distribution in plant height (cm) of progeny derived from initial mating of determinate (dede) x indeterminate (DeDe) cucumber (Cucumis sativus L.) inbreds. Plant heights in an F2 population (panel a), an F2 derived F3 population (panel b) segregating plants in an BC1 S2 population showing a range in determinate and indeterminate individuals classified in a BC1 S2 population (panel c) and in BC1 S2 plants derived from self pollination of BC1 S2 plants (panel d).
Literature Cited
- Staub, J.E., L.D. Knerr, and H.J. Hopen. 1992. Effects of plant density andherbicides on cucumber productivity. J. Amer. Soc. Hort. Sci. 117:48-53.