Cucurbit Genetics Cooperative Report 20:50-52 (article 22) 1997
Humberto Rios Labrada1 , Antonio Fernandez Almirall2 and Enrique Casanova Galarraga3
1 Instituto Nacional de Ciencia Agricola, INCA GP No.1 San jose de las Lajas, La Habana, Cuba, CP 32700
2 Instituto Superior Pedagogico para la ensenanza Tecnica y Profesional, Rpto Alday, La Habana, Cuba
3 Instituto Superior Pedagogico Juan Marinello, Matanzas, Cuba
Maynard (1996) recently reported on the promising potential for developing high-yielding pumpkin hybrids when grown under high-input cropping conditions. He showed that heterosis could be used as a straightforward means to increase pumpkin yield per area. On the contrary, under the low-input cultural conditions common in Cuba, a different breeding strategy must be used and pumpkin cultivars should be developed under conditions of stress and stress interactions interactions. That is why a collection of germplasm resistant or tolerant to marginal environments has been an essential issue for Cuban pumpkin breeders. The aim of this paper is to show the role of landraces as an important source of genetic material for tolerance to marginal growth conditions, and to show how a simple selection method was able to combine genes for a sustainable response to biotic and abiotic stress interactions under Cuban cultural conditions.
Materials and Methods. Nine selected lines were developed from Cuban landraces using recurrent selection with testing of open pollinated sib families. The lines were cultivated on farms providing latosolic soils and different abiotic and biotic stresses. Performance was determinated on the basis of yield as well as the occurrence of silvering leaves (Paris, 1987) and downy mildew infection (Ciba Geigy, 1984).
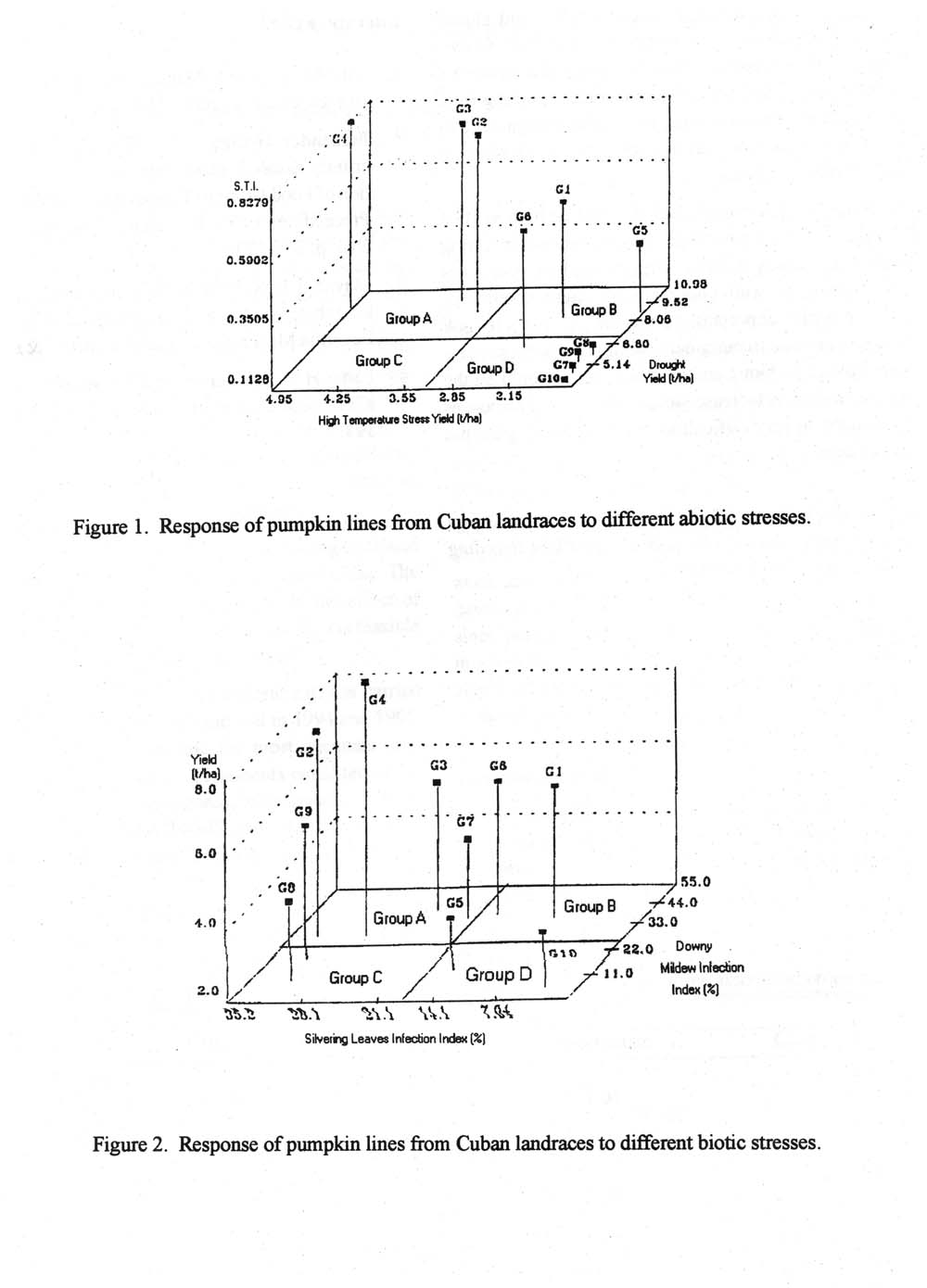
Taking into consideration Fernandez/s criteria (1993), the genotypes were classified according to their reaction to abiotic stresses as follows:
- Group A: Genotypes performing favorably under high temperature and drought stresses.
- Group B: Genotypes perform favorably under high temperature stress and poorly under drought stress.
- Group C: Genotypes performing poorly under high temperature stress and favorably under drought stress.
- Group D: Genotypes performing poorly under both abiotic stress conditions.
In addition, yield was plotted against genotype infection indexes for silvering leaves and downy mildew symptoms under naturally high disease conditions to characterize their biotic stress performance. Genotypes were classified according to these criteria as follows:
- Degree 1: Genotypes having a high infection index to downy mildew and silvering leaves (GROUP A).
- Degree 2: Genotypes having a high infection index to downy mildew and high infection index to silvering leaves (GROUP B).
- Degree 3: Genotypes having a low infection index to downy mildew and high infection index to silvering leaves (GROUP C).
- Degree 4: Genotypes having a low infection index for both diseases (GROUP D).
Results and Discussion. For abiotic stress (i.e., heat and drought), genotypes G2, G3, and G4 had a high value of stress tolerance and were included in Group A (Figure 1). Group B was represented by G1 and G5; the first genotype had a medium and the second a low stress tolerance index value. There were no genotypes included in the Group C cluster. Genotypes G6, G7, G8, G9, and G10 were included in the D cluster, characterized by low tolerance to heat and drought stresses. G10 has been marketed by a Cuban seed company nearly everywhere as a unique commercial variety.
Deviating performances were evident for genotypes G1 and G2, selected from landrace P-1523, and for genotypes G3 and G4, selected from landrace P-130. P-1523 was collected in The Isle of Youth and P-130 in San Juan y Martinez, Pinar del Rio. Both places posses dry soils and a rainfall regimen similar to the Cuban average. Thus, presumably, the different allelic series were linked and arose from natural selection to biotic and abiotic stress interactions under marginal conditions. Consequently, when breeding plants for marginal conditions it is important to test not only the landraces coming from there, but also those coming from other marginal environments. Evolution may have proceeded differently in the different locations.
Surprisingly, genotypes G2 and G4 had the highest yield (Figure 2) despite their high infection index to silvering leaves and downy mildew. Previously, both genotypes had performed well under abiotic stress conditions. Thus, it is also important in a breeding program to look for this response to marginal conditions, since a sustainable linkage to biotic and abiotic stresses would be the desired means to increase yields and to reveal responses that might be very difficult to observe under high-input agriculture.
The authors gratefully acknowledge the manuscript suggestions carried by Dr I. Boz from Plant Breeding Department of Wageningen.
Figure 1. Response of pumpkin lines from Cuban landraces to different abiotic stresses.
Figure 2. Response of pumpkin lines from Cuban landraces to different biotic stresses.
Literature Cited
- Ciba-Geigy. 1984. Manual para ensayos de campo en proteccion vegetal. CIBA_GEIC xii. 205p.
- Fermandez George, C.J. 1992. Effective selection criteria assesing plant stress tolerance. In: Adaptation of Food Crops to Temperature and Water Stress: Proceedings of an International Symposium. Taiwan. p. 257-270.
- Maynard, D.N. 1996. Performance of tropical pumpkin inbreds and hybrids in: Toward 2000: Proc. 6th Eucarpia Meeting on Cucurbit Genetics & Breeding.
- Paris, H S., H. Nerson and Y. Burger. 1987. Leaf silvering of Cucurbita.Canadian J. Science 67: 593-598.
The authors gratefully acknowledge the manuscript suggestions carried by Dr I. Boz from Plant Breeding Department of Wageningen.